|
|

Efecto del sustrato de oviposición en la eficiencia reproductiva del grillo Acheta domesticus (Orthoptera: Gryllidae)
Jennifer Canales-Cubillo1![]() , Jairo Mora-Prendas2
, Jairo Mora-Prendas2![]()
1. Universidad de Costa Rica, Sede Guanacaste, Recinto de Santa Cruz, Guanacaste, Costa Rica; jennifer.canalescubillo@ucr.ac.cr
2. Universidad de Costa Rica, Sede Guanacaste, Recinto de Santa Cruz. Laboratorio de Entomología, Guanacaste, Costa Rica; jairo.mora@ucr.ac.cr
Recibido 10-XI-2025 ● Corregido 23-III-2026 ● Aceptado 25-III-2026
DOI: https://doi.org/10.22458/urj.v18i1.6175
ABSTRACT. “Effect of oviposition substrate on the reproductive efficiency of the cricket Acheta domesticus (Orthoptera: Gryllidae)”. Introduction: The significance of the house cricket (Acheta domesticus) as a protein source for human and animal nutrition, coupled with the recent surge of the edible insect industry, highlights the need to develop and standardize reproductive protocols. These protocols must optimize oviposition, embryonic development, and species survival within intensive rearing systems. Objective: To evaluate the quality and preference of organic substrates as oviposition media for female A. domesticus. Methods: From October to December 2024, we tested four organic substrates—river sand, coconut fiber, agricultural soil, and peat moss—on A. domesticus reproduction through two trials. The first, a randomized design with four treatments and six replications to assess substrate quality. The second, an exploratory trial where all substrates were offered simultaneously to determine oviposition preference. Results: Peat moss was preferred and had the highest nymph emergence. Conclusion: Peat moss stood out as the most efficient substrate, presumably due to its moisture retention capacity, porosity, and thermal stability, while other substrates negatively affected embryonic development.
Keywords: Embryonic viability, humidity, development, hatching, incubation, insect, microorganisms
RESUMEN. Introducción: la importancia del grillo doméstico (Acheta domesticus) como fuente de proteína para la nutrición humana y animal, junto con el reciente auge de la industria de insectos comestibles, resalta la necesidad de desarrollar y estandarizar protocolos reproductivos. Estos protocolos deben optimizar la oviposición, el desarrollo embrionario y la supervivencia de la especie dentro de sistemas de cría intensiva. Objetivo: evaluar la calidad y preferencia de sustratos orgánicos como medios de oviposición para hembras de A. domesticus. Métodos: de octubre a diciembre de 2024 evaluamos cuatro sustratos orgánicos—arena de río, fibra de coco, suelo agrícola y turba—sobre la reproducción de A. domesticus con dos ensayos. El primero consistió en un diseño aleatorizado con cuatro tratamientos y seis repeticiones para evaluar la calidad del sustrato. El segundo fue un ensayo exploratorio en el que todos los sustratos se ofrecieron simultáneamente para determinar la preferencia de oviposición. Resultados: la turba fue el sustrato preferido y presentó la mayor emergencia de ninfas. Conclusión: la turba destacó como el sustrato más eficiente, presumiblemente debido a su capacidad de retención de humedad, porosidad y estabilidad térmica, mientras que los otros sustratos afectaron negativamente el desarrollo embrionario.
Palabras clave: viabilidad embrionaria, humedad, desarrollo, eclosión, incubación, insecto, microorganismos
Acheta domesticus, especie insectil destacada de la subfamilia Gryllinae, orden Orthoptera, presenta características morfológicas distintivas que facilitan su identificación, tales como una fila de espinas largas en las tibias posteriores, dos cercos sensoriales en la región terminal del abdomen, ocelos dispuestos en triángulo obtuso y un ovopositor igual o más alargado que la mitad de los fémures posteriores (Triplehorn & Johnson, 2005). La especie presenta un marcado dimorfismo sexual, en donde las hembras adultas son más robustas que los machos (Santos & Bernardo, 2005) y poseen un ovopositor prolongado en el extremo abdominal (Siddiqui et al., 2024; Triplehorn & Johnson, 2005) que les permite depositar los huevos en el sustrato (Cruz et al., 2021).
En condiciones de vida libre, las hembras de la subfamilia Gryllidae, depositan sus huevos en sustratos con altos contenidos de humedad (De Farias-Martins et al., 2017; Walker, 1999), lo que les proporciona un microambiente favorable para la incubación y el desarrollo embrionario (Mitchaothai et al., 2024). Tanto la humedad como la temperatura del sustrato influyen significativamente en la eclosión de los huevos, siendo ambas condiciones críticas para el proceso (Wu et al., 2021).
La especie fue incorporada en el continente americano para ser utilizada como alimento vivo para animales exóticos (Finke, 2015; Valdés et al., 2022) y como fuente complementaria de nutrientes en la alimentación humana (Farkas et al., 2025; Fleta Zaragozano, 2018; Rumpold & Schlüter, 2013; Siddiqui et al., 2024; Yi et al., 2013). Su cuerpo contiene todos los aminoácidos esenciales (Pilco-Romero et al., 2023; Udomsil et al., 2019) con un valor proteico del 67,66%, 4,73% de cenizas, 11,60% de fibra, 1,17% de grasas y 14,84% de carbohidratos (Morillo Cruz & Villegas Suclupe, 2024). Estos atributos posicionan a la especie como una de las más prometedoras para diversificar las fuentes proteicas.
En Costa Rica la industria de insectos comestibles ha experimentado un crecimiento acelerado tras la resolución N.º SENASA-DG-R0026-2021 que autorizó a A. domesticus como una de las tres especies aptas para su cría comercial (Servicio Nacional de Salud Animal, 2021). No obstante, este sector enfrenta desafíos relacionados con la falta de información técnica aplicada, la necesidad de estandarizar prácticas de manejo y la optimización de sistemas de producción que garanticen eficiencia y competitividad (Bermúdez-Serrano et al., 2023).
Para optimizar la cría comercial de A. domesticus, resulta fundamental generar un conocimiento detallado de su biología, dado que estos aspectos influyen directamente en su manejo y reproducción. En este contexto, nuestro estudio se enfoca en evaluar la calidad y la preferencia de los sustratos orgánicos como medio de oviposición, con el objetivo de aportar información que contribuya a mejorar la calidad y productividad de ninfas durante las fases iniciales de emprendimientos emergentes.
MATERIALES Y MÉTODOS
Llevamos a cabo la investigación durante el cuarto trimestre de 2024 (octubre-diciembre), en la granja de insectos de la Universidad de Costa Rica, ubicada en la Finca Experimental de Santa Cruz (FESC), Guanacaste. Dicha granja está localizada a 41 metros sobre el nivel del mar (41m s. n. m.) bajo las coordenadas geográficas 10°28’54” N y 85°59’28” O. El sitio de estudio presenta condiciones climáticas características de la zona guanacasteca con temperaturas promedio de 27,90°C (Cerdas-Ramírez, 2015) y una humedad relativa media del 74% (Vega Villalobos & Salas Camacho, 2012).
Realizamos dos ensayos independientes para evaluar, respectivamente, (1) la calidad del sustrato y (2) la preferencia de oviposición de adultos de A. domesticus (Orthoptera: Gryllidae), valorados en términos de postura y éxito de eclosión de ninfas. En ambos experimentos utilizamos cuatro tipos de sustrato como medios de oviposición: arena de río, fibra de coco, tierra agrícola y turba. Los ensayos se desarrollaron bajo condiciones ambientales no controladas, con una temperatura promedio de 37°C y una humedad relativa del 80%.
Antes de iniciar los ensayos, preparamos las unidades experimentales correspondientes a cada experimento. Todas fueron idénticas en cuanto al tamaño y a los elementos dispuestos en su interior, con la única diferencia del número de insectos introducidos. Cada unidad experimental consistió en una caja plástica con tapa y una capacidad de 70 litros (55cm de largo × 38cm de ancho × 33,5cm de alto). En la parte superior de los lados laterales de cada caja abrimos seis orificios (tres a cada lado), cada una con un diámetro de 55mm, los cuales cubrimos con malla antiáfidos para garantizar una ventilación adecuada. En su interior colocamos cuatro cartones de huevos como refugio, un bebedero para canarios de 60mL y un plato Petri con alimento balanceado para gallinas ponedoras.
Calidad de sustrato: en este primer ensayo utilizamos un diseño irrestricto al azar (DCA) con cuatro tratamientos y seis repeticiones, totalizando 24 unidades experimentales. En cada unidad experimental introdujimos diez hembras vírgenes y tres machos de A. domesticus. Los tratamientos se aplicaron individualmente en cada unidad experimental mediante frascos plásticos de 60mL con múltiples perforaciones en la tapa. Rellenamos totalmente el interior de cada frasco con un sustrato previamente humedecido (arena de río, fibra de coco, tierra agrícola o turba) y lo ubicamos dentro de la unidad correspondiente, según el tratamiento asignado.
Permitimos que las hembras ovipositaran durante un periodo de 168 horas (siete días) (Fig. 1 A). Al finalizar ese tiempo, retiramos cada frasco y trasvasamos su contenido a un recipiente plástico de mayor capacidad (946mL), en donde también colocamos un trozo de cartón de huevos (i.e., dos celdas) como refugio (Fig. 1 B). Etiquetamos cada recipiente con los datos del tratamiento, número de repetición y fecha. Los recipientes se sellaron con sus tapas originales, en las cuales se realizaron perforaciones para asegurar una ventilación adecuada. A partir de ese momento, revisamos el contenido de cada recipiente cada tres días para verificar la humedad del sustrato por medio de una inspección visual. Cuando fue necesario, humedecimos nuevamente el material para favorecer la eclosión de ninfas. Tan pronto como observamos ninfas emergidas, retiramos momentáneamente los cartones, los sacudimos dentro de frascos de 946mL y recolectamos las ninfas con ayuda de un aspirador entomológico. Conservamos los ejemplares en alcohol al 95% para su posterior conteo (Fig. 1 C). Repetimos este procedimiento durante siete días consecutivos para asegurar la recolección del mayor número posible de ninfas.

Fig. 1. Ensayos de calidad y preferencia de sustratos para la oviposición de A. domesticus. A. Oviposición de hembras, B. Emergencia y cosecha de ninfas, C. Almacenamiento de ninfas en alcohol.
Preferencia de sustrato: a diferencia del primer ensayo, en este no aplicamos un diseño estadístico definido; sin embargo, incrementamos el número de unidades experimentales a 15, con el fin de obtener datos más concluyentes. En cada unidad incorporamos inicialmente quince hembras y cinco machos de A. domesticus, cuya descendencia generó aproximadamente 300 parejas reproductoras y con esta población iniciamos el experimento.
Basamos el ensayo en un modelo de libre elección, en el cual, las 300 parejas reproductivas fueron expuestas simultáneamente a los cuatro sustratos en evaluación: arena de río, fibra de coco, tierra agrícola y turba. Los materiales se dispusieron aleatoriamente dentro de cada recinto utilizando frascos de 60 mL con tapas perforadas. Esta configuración facilitó la identificación de sus preferencias conductuales de oviposición bajo condiciones de competencia real. Una vez concluida la fase de exposición se replicó el protocolo de obtención de resultados y procesamiento de datos detallado en el ensayo anterior.
Recolección y análisis de datos: posteriormente a la eclosión de las ninfas las almacenamos en viales de vidrio con alcohol al 95%. Luego las cuantificamos utilizando un estereoscopio modelo NexiusZoom, pinzas entomológicas y viales de clasificación. Registramos los datos en una hoja de Excel® y, a partir de esta, generamos las matrices necesarias para el análisis. Iniciamos con la prueba de normalidad de Shapiro-Wilk, al no cumplirse este supuesto empleamos la prueba no paramétrica de Kruskal-Wallis. Posteriormente, realizamos comparaciones múltiples mediante la prueba de Dunn con corrección de Bonferroni, con el objetivo de identificar diferencias significativas entre tratamientos. Todos los análisis fueron realizados en el software R (versión 4.5.1; R Core Team, 2025).
RESULTADOS
Encontramos diferencias significativas en el ensayo de calidad de los sustratos entre los tratamientos (p= 8.14 × 10⁻⁴; Kruskal-Wallis). La mayor emergencia de ninfas la obtuvimos con la turba (n= 1917 ± 139), con una significativamente mayor a los demás tratamientos: tierra agrícola (n= 787 ± 133; p = 1,04 × 10⁻²), fibra de coco (n= 719 ± 50; p = 6,7 × 10⁻⁶) y arena de río (n= 0; p= 1,0 × 10⁻⁷) (Fig. 2). A pesar de que los porcentajes de eclosión en tierra agrícola (23%) y fibra de coco (21%) fueron inferiores al registrado en la turba (56%, n = 3423), ambos sustratos superaron al tratamiento con arena de río, en donde no observamos emergencia de individuos.
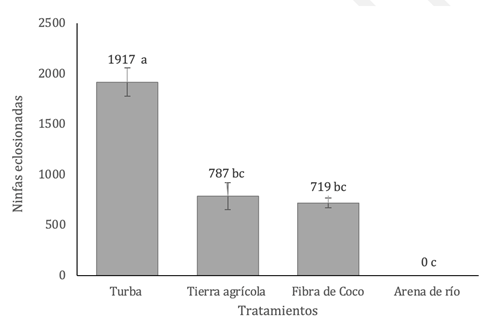
Fig. 2. Emergencia de ninfas de A. domesticus bajo diferentes sustratos de oviposición, en el total de unidades experimentales. Letras distintas indican diferencias significativas (p≤0,05), según Dunn con corrección de Bonferroni, tras Kruskal-Wallis.
La turba se posicionó como el sustrato más eficiente para la emergencia de ninfas, al presentar la mayor media (X̅ = 320) y un coeficiente de variación relativamente bajo (CV = 43%). En segundo lugar, se ubicó la fibra de coco con una emergencia moderada y una variabilidad aceptable entre réplicas. En contraste, la tierra agrícola evidenció una alta dispersión en los datos (CV = 101%), lo que comprometió su confiabilidad como sustrato de oviposición. (TABLA 1)
TABLA 1
Número de ninfas de A. domesticus emergidas en función del sustrato de oviposición, por unidad
experimental
|
Ninfas emergidas |
Media |
Mediana |
Ámbito |
Desviación estándar |
CV% |
|
|
Arena de río |
0 |
0 |
0 |
0 |
0 |
0 |
|
Fibra de coco |
719 |
120 |
105 |
153 |
50 |
42 |
|
Tierra agrícola |
787 |
131 |
58 |
316 |
133 |
101 |
|
Turba |
1 917 |
320 |
349 |
374 |
139 |
43 |
Para el análisis de la preferencia de sustratos encontramos diferencias significativas entre los tratamientos evaluados (p = 9.45 × 10⁻⁴; Kruskal-Wallis). La turba presentó la mayor eclosión de ninfas (n= 15 177 ± 445) con valores significativamente superiores a los observados en tierra agrícola (n= 3 469 ± 228; p = 3,4×10⁻⁴), fibra de coco (n= 1 557 ± 140; p= 4,5×10⁻⁵) y arena de río (n= 219 ± 16; p = 1,9×10⁻⁵). Por otro lado, los tratamientos de tierra agrícola y fibra de coco no difirieron entre sí, aunque la emergencia de ninfas en ambos sustratos fue significativamente mayor que la observada en la arena de río (Fig. 3).
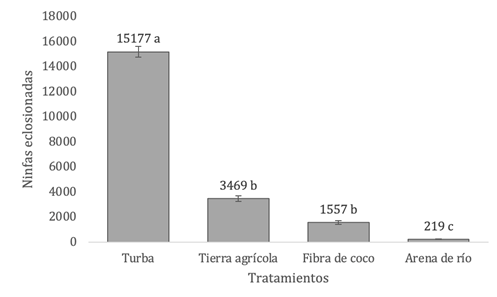
Fig. 3. Emergencia de ninfas de A. domesticus bajo diferentes sustratos de oviposición, en el total de las unidades experimentales. Letras distintas indican diferencias significativas (p≤0,05), según Dunn con corrección de Bonferroni, tras Kruskal-Wallis.
La turba se posicionó nuevamente como el mejor sustrato de oviposición con la media más alta de nacimientos (X̅= 1012) y el menor coeficiente de variación (CV= 44%). En segundo lugar, se ubicaron la tierra agrícola y la fibra de coco, las cuales mostraron una emergencia moderada y una variabilidad aceptable entre réplicas. En contraste, la arena de río presentó la menor eclosión de ninfas (=15). (TABLA 2)
TABLA 2
Preferencia de sustratos de oviposición por parte de las hembras de A. domesticus medido en términos de ninfas emergidas
|
Tratamiento |
Ninfas emergidas |
Media |
Mediana |
Ámbito |
Desviación estándar |
CV% |
|
Arena de río |
219 |
15 |
10 |
56 |
16 |
112 |
|
Fibra de coco |
1557 |
104 |
53 |
563 |
140 |
134 |
|
Tierra agrícola |
3469 |
231 |
177 |
792 |
228 |
99 |
|
15 177 |
1012 |
949 |
1558 |
445 |
44 |
* Los valores presentados representan la emergencia de ninfas obtenida en cada tratamiento, calculada a partir de 15 repeticiones por tratamiento.
Las propiedades inherentes y estructurales de un sustrato son cruciales para el desarrollo embrionario de los huevos en múltiples especies animales, tales como reptiles e insectos (Bell et al., 2024; Packard et al., 1987; Wu et al., 2021). Estudios realizados por Tanga et al. (2015) demostraron que los sustratos influyen significativamente en el comportamiento de oviposición de las hembras de A. domesticus, las cuales seleccionan el medio para depositar sus huevos en función de sus características físicas y químicas. Asimismo, Kobe Solares & Murillo-Hiller (2021) señalan que uno de los principales atributos que debe cumplir un sustrato para favorecer una adecuada oviposición y eclosión de ninfas es su contenido de humedad, esta debe mantenerse en un rango óptimo entre el 50 y 60%. En particular, un mayor contenido de humedad en el sustrato se asocia con un aumento en la oviposición (De Farias-Martins et al., 2017), mientras que su temperatura afecta tanto la duración del desarrollo como la tasa de eclosión de los huevos en condiciones naturales (Odhiambo et al., 2022; Wu et al., 2021).
En esta investigación la turba promovió la mayor eclosión de ninfas en ambos ensayos realizados, alcanzando niveles significativamente superiores en comparación con los demás sustratos evaluados. Este comportamiento podría atribuirse a las propiedades intrínsecas del sustrato como la alta capacidad de retención de humedad, baja densidad aparente, elevada porosidad y adecuada regulación térmica (Kitir et al., 2018). En contraste con otros sustratos, como la fibra de coco o la tierra agrícola, la turba presenta una mayor uniformidad en sus características físicas, lo que podría explicar la preferencia marcada de las hembras por este material en ambos ensayos.
Si bien, la fibra de coco y la tierra agrícola comparten características físicas similares, como la capacidad de retención de agua y la porosidad (Pire & Pereira, 2003), pueden presentar variaciones externas que comprometen su calidad como sustrato. En el caso de la fibra de coco, el método de procesamiento varía según la casa comercial que la produzca, lo cual puede modificar parámetros como la acidez y la salinidad del material. Estos cambios no solo afectan el proceso de oviposición, sino que también pueden favorecer la aparición de plagas y enfermedades (Puerta, 2012; Quesada, 2005; Soto Bravo & Betancurt Flores, 2022). De forma similar, la tierra agrícola presenta variaciones en su estructura y composición química según el sitio de procedencia; por ejemplo, en este ensayo se utilizó un suelo de textura arcillosa con características vérticas típicas de los Vertisoles de la bajura guanacasteca (Garbanzo-León et al., 2021). Este tipo de suelo tiende a presentar limitaciones físicas como baja porosidad y escasa estabilidad estructural, debido a que al humedecerse adquiere una consistencia pegajosa y al secarse se compacta con facilidad (Cabidoche & Ruy, 2001; Teshale, 2023), lo que puede restringir el intercambio gaseoso en los embriones en formación y dificultar la oviposición por parte de las hembras.
En cuanto al sustrato de arena de río, sus características físicas, como la baja retención de agua y la textura granulada (Garbanzo-León et al., 2021), sumada a las altas temperaturas registradas en Santa Cruz, Guanacaste, pudieron influir en la baja eclosión durante los dos ensayos. Diversos estudios demuestran que las hembras prefieren ovipositar en sustratos con mayor contenido de humedad (Destephano et al., 1982; De Farias-Martins et al., 2017; Tanga et al., 2015). En este sentido, Kobe Solares y Murillo Hiller (2021) señalan que el rango óptimo de humedad para asegurar una eclosión exitosa se sitúa entre el 50 y el 60%. Durante la etapa reproductiva las hembras experimentan una serie de conductas y cambios físico-sensoriales que les permiten determinar las condiciones y ambientes óptimos para la oviposición. De Farias-Martins et al. (2017), Shields et al. (2022) y Tanga et al. (2015) indicaron que los insectos desarrollan en sus antenas higrorreceptores (detectores de humedad) y quimiorreceptores (olfativos y gustativos), los cuales les permiten identificar los ambientes más favorables y seleccionar el sustrato con el balance hídrico adecuado para depositar sus huevos. Partiendo de lo anterior, las propiedades de la arena pueden hacerla menos atractiva como sustrato de oviposición al no proporcionar los estímulos sensoriales ni la humedad necesaria. Además, al ser un sustrato de origen fluvial, presenta mayor vulnerabilidad a la proliferación de microorganismos patógenos como el moho verde, capaces de destruir los huevos (Candan, 2018). Por estas razones, la arena de río se considera un sustrato no recomendable para este tipo de experimentos o en su defecto realizar algún método de esterilización para evitar dichos problemas.
Los resultados del estudio confirman que las propiedades fisicoquímicas del sustrato son fundamentales para garantizar el éxito reproductivo y la viabilidad embrionaria de A. domesticus. La turba se destacó como el sustrato más eficiente y preferido por las hembras, presuntamente por su alta retención de humedad, porosidad y estabilidad térmica, mientras que la fibra de coco, la tierra agrícola y la arena de río presentaron limitaciones físicas que comprometieron la oxigenación y la integridad de los huevos. Por tanto, seleccionar cuidadosamente el sustrato de oviposición, priorizando su capacidad hídrica, estabilidad térmica e inocuidad microbiológica, resulta clave para optimizar la reproducción y la supervivencia embrionaria en sistemas de cría controlada de A. domesticus.
AGRADECIMIENTOS
Agradecemos a la Universidad de Costa Rica y a la Vicerrectoría de Investigación por financiar el proyecto principal que dio origen a esta investigación: C3452 “Cría y producción de grillos (Acheta domesticus) como fuente de proteína y otros minerales destinados a la alimentación animal, en el trópico seco de Costa Rica”.
ÉTICA, CONFLICTO DE INTERESES Y DECLARACIÓN DE FINANCIAMIENTO
Los autores declaramos haber cumplido plenamente con todos los requisitos éticos y legales aplicables durante el desarrollo del estudio y la elaboración del manuscrito. Igualmente, confirmamos que no existen conflictos de interés de ningún tipo, que todas las fuentes de financiamiento se encuentran completa y claramente indicadas en la sección de agradecimientos y que estamos totalmente de acuerdo con la versión final editada del artículo. Se ha conservado un documento firmado en los archivos de la revista.
La declaración de la contribución de cada autor al manuscrito es la siguiente: J.C.C. y J.M.P.: diseño del estudio, recopilación y análisis de datos. J.C.C.: Recogida de datos. Todos los coautores: preparación y aprobación final del manuscrito.
REFERENCIAS
Bell, C., Raynal, R. S., Noble, D. W. A., Schwanz, L. E., Warner, D. A., Pruett, J. E., & Riley, J. L. (2024). The effect of moisture during development on phenotypes of egg-laying reptiles: A systematic review and meta-analysis. Journal of Experimental Biology, 228(4), JEB249960. https://doi.org/10.1242/jeb.249960
Bermúdez-Serrano, I. M., Quirós-Blanco, A. M., & Acosta-Montoya, Ó. (2023). Mercado, oportunidades y retos de la producción, procesamiento y comercialización de insectos comestibles en Costa Rica y a nivel internacional. Agronomía Mesoamericana, 34(3), 53052. https://doi.org/10.15517/am.2023.53052
Cabidoche, Y.-M., & Ruy, S. (2001). Field shrinkage curves of a swelling clay soil: Analysis of multiple structural swelling and shrinkage phases in the prisms of a Vertisol. Australian Journal of Soil Research, 39(1), 143–160. https://doi.org/10.1071/SR99132
Candan, E. D. (2018). Molecular identification of fungal isolates and hatching success of green turtle (Chelonia mydas) nests. Archives of Microbiology, 200(6), 911–919. https://doi.org/10.1007/s00203-018-1496-0
Cerdas-Ramírez, R. (2015). Comportamiento productivo del pasto maralfalfa (Pennisetum sp.) con varias dosis de fertilización nitrogenada. InterSedes, 16(33), 125–145.
Cruz, D., Arévalo, H., & Vernot, D. (2021). Artrópodos: Producción de grillos de forma sustentable. Universidad de La Sabana; Minciencias; Gobernación de Cundinamarca; Arthro Food.https://doi.org/10.5294/978-958-12-0594-3
De Farias-Martins, F., Sperber, C. F., Albeny-Simões, D., Breaux, J. A., Fianco, M., & Szinwelski, N. (2017). Forest litter crickets prefer higher substrate moisture for oviposition: Evidence from field and lab experiments. PLoS ONE, 12(10), e0185800. https://doi.org/10.1371/journal.pone.0185800
Destephano, D. B., Brady, U. E., & Farr, C. A. (1982). Factors influencing oviposition behavior in the cricket, Acheta domesticus. Annals of the Entomological Society of America, 75(2), 111–114. https://doi.org/10.1093/aesa/75.2.111
Farkas, V. I., Máté, M., Takács, K., & Jánosi, A. (2025). The house cricket (Acheta domesticus Linnaeus) in food industry: Farming, technological challenges, and sustainability considerations. Applied Sciences, 15(17), 9494. https://doi.org/10.3390/app15179494
Finke, M. D. (2015). Complete nutrient content of four species of commercially available feeder insects fed enhanced diets during growth. Zoo Biology, 34(6), 554–564. https://doi.org/10.1002/zoo.21246
Fleta Zaragozano, J. (2018). Entomofagia: ¿Una alternativa a nuestra dieta tradicional? Sanidad Militar, 74(1), 41–46. https://doi.org/10.4321/S1887-85712018000100008
Garbanzo-León, G., Alvarado-Hernández, A., Vargas-Rojas, J. C., Cabalceta-Aguilar, G., & Vega-Villalobos, E. V. (2021). Nitrogen and potassium fertilization in corn in an Alfisol of Guanacaste, Costa Rica. Agronomía Mesoamericana, 32(1), 137–148. https://doi.org/10.15517/am.v32i1.39822
Kitir, N., Yildirim, E., Şahin, Ü., Turan, M., Ekinci, M., Ors, S., Kul, R., Ünlü, H., & Ünlü, H. (2018, November 5). Peat use in horticulture. In Peat [Peer-reviewed chapter]. IntechOpen. https://doi.org/10.5772/intechopen.79171
Kobe Solares, A., & Murillo Hiller, L. R. (2021). Potencial invasor del grillo eurasiático Acheta domesticus (Orthoptera: Gryllidae) en Costa Rica. Cuadernos de Investigación UNED, 13(2).http://dx.doi.org/10.22458/urj.v13i2.3259
Mitchaothai, J., Lertpatarakomol, R., Trairatapiwan, T., & Lukkananukool, A. (2024). Influence of incubation temperature and relative humidity on the egg hatchability pattern of two-spotted (Gryllus bimaculatus) and house (Acheta domesticus) crickets. Animals, 14(15), 2176. https://doi.org/10.3390/ani14152176
Morillo Cruz, C. G., & Villegas Suclupe, P. R. (2024). Potencial proteico de harina de grillo (Acheta domesticus) como una alternativa sostenible para el consumo humano. Revista Científica Pakamuros, 12(1), 16–26. https://doi.org/10.37787/jh8b0c66
Odhiambo, M. A., Olweny, C. O., & Okuto, E. O. (2022, September). Effects of temperature on the development and survival of cricket species: Acheta domesticus and Gryllus bimaculatus (Orthoptera: Gryllidae). East African Journal of Agriculture and Biotechnology, 5(1), 176–189. https://doi.org/10.37284/eajab.5.1.834
Packard, G., Packard, M. J., & Miller, K. (1987). Influence of moisture, temperature, and substrate on snapping turtle eggs and embryos. Ecology, 68(4), 983–993. https://doi.org/10.2307/1938369
Pilco-Romero, G., Chisaguano-Tonato, A. M., Herrera-Fontana, M. E., Chimbo-Gándara, L. F., Sharifi-Rad, M., Giampieri, F., Battino, M., Vernaza, M. G., & Álvarez-Suárez, J. M. (2023). House cricket (Acheta domesticus): A review based on its nutritional composition, quality, and potential uses in the food industry. Trends in Food Science & Technology, 142, 104226. https://doi.org/10.1016/j.tifs.2023.104226
Pire, R., & Pereira, A. (2003). Propiedades físicas de componentes de sustratos de uso común en la horticultura del estado Lara, Venezuela: Propuesta metodológica. Bioagro, 15(1), 55–64. http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S1316-33612003000100007
Puerta, C. E. (2012). Producción de plántulas de pimentón (Capsicum annuum L.) en sustratos orgánicos a base de mezclas con fibra de coco. Revista Científica UDO Agrícola, 12(2), 298–306. https://dialnet.unirioja.es/servlet/articulo?codigo=4688395&utm
Quesada, G. M. C. (2005). Evaluación de sustratos para almácigos de hortalizas. Agronomía Mesoamericana, 16(2), 171–183. https://www.redalyc.org/pdf/437/43716207.pdf
R Core Team. (2025). R: A language and environment for statistical computing. The R Foundation for Statistical Computing. https://www.R-project.org
Rumpold, B. A., & Schlüter, O. K. (2013). Nutritional composition and safety aspects of edible insects. Molecular Nutrition & Food Research, 57(5), 802–823. https://doi.org/10.1002/mnfr.201200735
Santos, C., & Bernardo, J. (2005). The quantitative genetic basis of female and male body size and their implications on the evolution of body size dimorphism in the house cricket Acheta domesticus (Gryllidae). Genética Evolutiva • Genet. Mol. Biol., 28(4). https://doi.org/10.1590/S1415-47572005000500030
Siddiqui, S. A., Zhao, T., Fitriani, A., Rahmadhia, S. N., Alirezalu, K., & Fernando, I. (2024). Acheta domesticus (house cricket) as human foods - An approval of the European Commission: A systematic review. Food Frontiers, 5(1), e358. https://doi.org/10.1002/fft2.358
Servicio Nacional de Salud Animal. (2021). Lista oficial de los nombres científicos de los animales considerados como domésticos, que se regularán por lo dispuesto en la Ley General del Servicio Nacional de Salud Animal, N.º 8495 y serán competencia exclusiva de esta Autoridad Sanitaria (Resolución SENASA-DG-R0026-2021). Ministerio de Agricultura y Ganadería. https://www.senasa.go.cr
Shields, V. D., Weaver, C. E., & Farmer, S. (2022). Functional morphology of sensory organs of the house cricket, Acheta domesticus (L.): A morphological and physiological investigation. The FASEB Journal, 36(S1). https://doi.org/10.1096/fasebj.2022.36.S1.R6108
Soto Bravo, F., & Betancurt Flores, A. (2022). Evaluación de metodologías para determinar las características físicas de un sustrato de fibra. Agronomía Costarricense, 46(2), 29–44. https://doi.org/10.15517/rac.v46i2.52044
Tanga, C., Akutse, K. S., Fiaboe, K. K. M., Niassy, S., Mohamed, S. A., & Ekesi, S. (2015, October). Effect of colour and substrate on oviposition behaviour and egg hatchability in the house cricket Acheta domesticus (L.) (Orthoptera: Gryllidae). Paper presented at the 21st Biannual Meeting and Scientific Conference, African Association of Insect Scientists (AAIS 2015), Cotonou, Benin. https://www.researchgate.net/publication/284109456
Teshale, E. (2023). Problems associated with physical and chemical properties of Vertisols and management options in agriculture: A review. Science Research, 11(5), 97–103. https://doi.org/10.11648/j.sr.20231105.11
Triplehorn, C. & Johnson, N. (2005). Borror and Delong’s Introduction to the Study of Insects. United States of America: Thomson-Brooks/Cole.
Udomsil, N., Imsoonthornruksa, S., Gosalawit, C., & Ketudat-Cairns, M. (2019). Nutritional values and functional properties of house cricket (Acheta domesticus) and field cricket (Gryllus bimaculatus). Food Science and Technology Research, 25(4), 597–605. https://doi.org/10.3136/fstr.25.597
Valdés, F., Villanueva, V., Durán, E., Campos, F., Avendaño, C., Sánchez, M., Domínguez-Araujo, C., & Valenzuela, C. (2022). Insects as feed for companion and exotic pets: A current trend. Animals (Basel), 12(11), 1450. https://doi.org/10.3390/ani12111450
Vega Villalobos, E. V., & Salas Camacho, R. E. (2012). Curvas de absorción de nutrientes bajo dos métodos de fertilización en sandía, en Guanacaste, Costa Rica. InterSedes: Revista de las Sedes Regionales, 13(26), 19–44.
Walker, T. J. (1999). House cricket, Acheta domesticus (Linnaeus). University of Florida, Institute of Food and Agricultural Sciences. https://edis.ifas.ufl.edu/publication/IN220
Wu, T., Hao, S., & Kang, L. (2021). Effects of soil temperature and moisture on the development and survival of grasshopper eggs in Inner Mongolian grasslands. Frontiers in Ecology and Evolution, 9, 727911. https://doi.org/10.3389/fevo.2021.727911
Yi, L., Lakemond, C. M. M., Sagis, L. M. C., Eisner-Schadler, V. R., van Huis, A., & van Boekel, M. A. J. S. (2013). Extraction and characterization of protein fractions from five insect species. Food Chemistry, 141(4), 3341–3348. https://doi.org/10.1016/j.foodchem.2013.05.115