|
|

Abundancia y riqueza de la subfamilia Cryptinae (Hymenoptera: Ichneumonidae) en un bosque tropical de Panamá
Jairo Mora-Prendas1![]()
1Universidad de Costa Rica, Laboratorio de Entomología, Sede Guanacaste, Recinto de Santa Cruz. Guanacaste, Costa Rica; jairo.mora@ucr.ac.cr
Recibido 27-I-2025 · Corregido 26-V-2025 · Aceptado 18-VI-2025
DOI: https://doi.org/10.22458/urj.v17i1.5658
ABSTRACT. “Abundance and richness of the subfamily Cryptinae (Hymenoptera: Ichneumonidae) in a tropical forest of Panama”. Introduction: Cryptinae is the most diverse and abundant subfamily within the Ichneumonidae. However, its study in Panama has focused on a limited number of species, hindering the understanding of its population dynamics in the region. Objective: To evaluate the abundance and species richness of Cryptinae wasps in a tropical forest of Panama. Methods: I analyzed 2 080 Ichneumonidae (Cryptinae) samples from Malaise traps on Barro Colorado Island, Panama, from the period 2014–2017. Results: Cryptinae were abundant but low in species richness, likely due to the island’s low elevation. Abundance was higher during the dry season and declined strongly in wetter years. Conclusion: Cryptinae is the most abundant Ichneumonidae subfamily in the island, with population patterns shaped by seasonal and climatic factors, particularly temperature and precipitation.
Keywords: wasps, parasitoids, seasonality, Hymenoptera, biological regulators, climate change.
RESUMEN. Introducción: Cryptinae es la subfamilia más diversa y abundante dentro de Ichneumonidae. Sin embargo, su estudio en Panamá se ha centrado en un número limitado de especies, lo que dificulta la comprensión de su dinámica poblacional en la región. Objetivo: Evaluar la abundancia y riqueza de especies de avispas Cryptinae en un bosque tropical de Panamá. Métodos: Analicé 2,080 muestras de Ichneumonidae (Cryptinae) recolectadas con trampas Malaise en la Isla Barro Colorado, Panamá, durante el período 2014–2017. Resultados: Cryptinae fue abundante, pero presentó baja riqueza de especies, probablemente debido a la baja altitud de la isla. La abundancia fue mayor durante la estación seca y disminuyó considerablemente en los años lluviosos. Conclusión: Cryptinae es la subfamilia más abundante de Ichneumonidae en la isla, con patrones poblacionales influenciados por factores estacionales y climáticos, especialmente la temperatura y la lluvia.
Palabras clave: avispas, parasitoides, estacionalidad, Hymenoptera, reguladores biológicos, cambio climático.
INTRODUCCIÓN
Los ichneumónidos son avispas parasitoides ampliamente distribuidas a nivel mundial (Ruíz-Cancino et al., 2012), responsables de regular las poblaciones de otros insectos en los ecosistemas naturales (Ruíz-Cancino & Kasparyan, 2000). Sus hábitos parasíticos, asociados con su amplia distribución geográfica, han convertido a este grupo de avispas en indicadores de la salud del ecosistema, ya que su éxito reproductivo está directamente relacionado con la diversidad y abundancia de los hospederos que atacan (Ruíz-Cancino et al., 2014).
Según la taxonomía de la familia Ichneumonidae, esta se clasifica en 42 subfamilias, que incluyen un total de 23 765 especies y 1 547 géneros descritos (Yu, DSK, 2024). En la región neotropical, la familia está representada por 31 de estas subfamilias (Hanson & Gauld, 2006) y un estimado de 7 400 especies (Ruíz-Cancino, 2015).
Dentro de Ichneumonidae, la subfamilia Cryptinae sobresale por ser una de las más abundantes y diversas, con al menos 4 500 especies descritas en todo el mundo (Hanson & Gauld, 2006). No obstante, su estudio en regiones como Panamá ha sido limitado, con registros centrados en unas pocas especies dentro de la subfamilia, dejando por fuera su biología y comportamiento reproductivo (Fincke, Higgins, & Rojas, 1990; Miranda et al., 2020; Murgas et al., 2021). El único estudio que aborda parcialmente estos aspectos es el realizado por Shapiro y Pickering (2000), quienes confirman que la abundancia de Cryptinae supera a la de otras 15 subfamilias en Barro Colorado.
Desde una perspectiva ecológica, Cryptinae cumple un rol clave como agente natural en la regulación de poblaciones de otros artrópodos, particularmente mediante el parasitismo de diversos estados de desarrollo (Miranda et al., 2020; Murgas et al., 2021; Álvarez-Cabrera et al., 2018). Estas avispas actúan como parasitoides idiobiontes (Mora-Prendas, 2025), desarrollándose sobre o dentro de sus hospedadores, los cuales suelen ser larvas o pupas de órdenes como Lepidoptera y Coleoptera (Hanson et al. 2006; Shapiro et al. 2000; Murgas et al., 2015). Mediante este mecanismo, las especies de Cryptinae contribuyen significativamente a la estabilidad ecológica y al control biológico natural, limitando el crecimiento poblacional de insectos fitófagos que, en condiciones de desequilibrio, pueden convertirse en plagas agrícolas o forestales.
A nivel global, la diversidad y distribución de Cryptinae han sido documentadas en diversos países. Por ejemplo, en un estudio realizado en Trabzon, Turquía, se registraron 26 muestras pertenecientes a 10 géneros y 10 especies, de las cuales dos fueron nuevos registros para la fauna turca (Çoruh & Kolarov, 2022). En el neotrópico, Santos y Hoppe (2018) identificaron 175 especies de Cryptinae distribuidas en 24 países a partir de 20 colecciones biológicas. En México, Cryptinae destaca como la subfamilia con la mayor cantidad de géneros y especies registradas (Ruíz-Cancino et al., 2014). Su relevancia es tal que, en la mayoría de los estudios, sobresale de entre otras subfamilias por su alta abundancia (Rodríguez et al., 2015; Álvarez et al., 2017; Pérez et al., 2018; Reátegui, 2019) y riqueza (Ruíz-Cancino, 2010; Ruíz-Cancino et al., 2011; Ruíz-Cancino et al., 2014; Ruíz-Cancino, 2015; González-Moreno et al., 2015). De manera similar, en Chile, registra 23 géneros de Cryptinae (Pádua et al, 2024), lo que refleja la gran diversidad del grupo.
En este contexto, resulta evidente la importancia de llevar a cabo investigaciones más detalladas sobre la subfamilia Cryptinae en diversas regiones del mundo. Estas investigaciones permitirían ampliar el conocimiento sobre su distribución, riqueza y ecología, por ejemplo, analizar sus patrones estacionales y la influencia del clima en las dinámicas de sus poblaciones. Este trabajo tiene como objetivo generar información básica y actualizada sobre las comunidades de Cryptinae de Barro Colorado, que sirva de base para futuros estudios de biodiversidad y conservación faunística en la región.
MATERIALES Y MÉTODOS
Analicé cuatro años de recolección de insectos (2014-2017), provenientes del proyecto mundial de estudio de bosques tropicales ForestGeo® (http://www.forestgeo.si.edu/), manejado por personal del Instituto de Investigaciones Tropicales Smithsonian en Isla Barro Colorado, Ciudad de Panamá. En el marco del proyecto, se instalaron 10 trampas Malaise de forma permanente en distintas secciones de la isla, con capturas semanales iniciadas desde el año 2000.
Las muestras correspondientes a los años seleccionados fueron trasladadas a las instalaciones del Programa Centroamericano de Maestría en Entomología (PCMENT) de la Universidad de Panamá para su procesamiento. En total, revisé 2 080 muestras de Hymenoptera con el objetivo de separar la familia Ichneumonidae de otros grupos de insectos. Posteriormente, clasifiqué la familia a nivel de subfamilias y morfoespecies, mediante el análisis de su morfología externa y el uso de claves taxonómicas (Hanson y Gauld, 2006). Con el fin de disminuir el sesgo por dimorfismo sexual en la determinación de las morfoespecies, realicé esta clasificación con base en las hembras. Posteriormente, asigné a los machos con mayores similitudes a cada morfoespecie. Con base en esta clasificación, analicé la abundancia, riqueza y distribución temporal de la subfamilia Cryptinae en Barro Colorado.
Con respecto a los datos meteorológicos utilizados en este estudio, los obtuve del Instituto de Investigaciones Tropicales Smithsonian, estos incluyeron registros de temperatura (°C), precipitación (mm) y humedad relativa (%), comprendidos entre los años 2014 al 2017. Las mediciones fueron realizadas con sensores especializados cada 10 minutos para temperatura y humedad, y cubos de inflexión para precipitación diaria. A partir de ellos, elaboré una base de datos en Excel con promedios mensuales y anuales. Adicionalmente, subdividí el año meteorológico en dos temporadas climáticas, de acuerdo con lo establecido por Eisenmann (1952) y Barone (1998). La temporada seca con una duración aproximada de 5 a 6 meses, que van de diciembre a mayo, mientras que la temporada lluviosa abarca los meses de junio a noviembre (6 y 7 meses).
Analicé la normalidad de los datos mediante el test de Shapiro-Wilk. Dado que la normalidad fue rechazada, apliqué pruebas no paramétricas: Wilcoxon para comparar la abundancia de dos temporadas (seca y lluviosa) y Kruskal-Wallis para tres o más comparaciones (años y temporadas por año). Así mismo, cuando fue necesario, se aplicaron las pruebas post hoc de Wilcoxon con corrección de Bonferroni. Los análisis se realizaron en R 3.4.0 (R Core Team, 2017), y se elaboraron tablas en Excel® para representar la abundancia y riqueza de Cryptinae por temporada y año.
RESULTADOS
Del procesamiento de 2 080 muestras de Hymenoptera recolectadas durante el periodo de estudio, obtuve en total 3 299 especímenes de la subfamilia Cryptinae. Este grupo presentó su mayor abundancia en el año 2015, seguido por 2014 y 2016, mientras que la menor abundancia se registró en 2017 (Tabla 1).
Estadística descriptiva de la subfamilia Cryptinae entre los años 2014 y 2017
|
Año |
Abundancia |
Promedio |
Media |
Ámbito |
Desviación estándar |
Coeficiente de variación (%) |
|
2014 |
830 |
69 |
46 |
149 |
48,3 |
70 |
|
2015 |
1171 |
98 |
94 |
164 |
50,1 |
51 |
|
2016 |
718 |
60 |
57 |
88 |
27,4 |
46 |
|
2017 |
580 |
48 |
45 |
86 |
26,5 |
55 |
|
Total |
3299 |
69 |
50 |
177 |
43,6 |
63 |
Al realizar el análisis individual de los años por temporadas climáticas (seca y lluviosa) (FIG. 1), el año 2014 presentó diferencia significativa entre ambas temporadas (W = 33, p = 0,0200), con una mayor abundancia en la época seca. De manera similar, el año 2015 presentó el mismo comportamiento (W = 35, p = 0,008).
En términos generales, se registró una abundancia significativamente mayor de Cryptinae durante la temporada seca en comparación con la lluviosa (Tabla 2.) (p = 0,002, prueba de Wilcoxon). El análisis de las temporadas específicas de cada año de recolección reveló que las temporadas lluviosas no presentaron diferencias significativas entre sí (p = 0,3176). En contraste, las temporadas secas mostraron diferencias significativas (p = 0,007), con variaciones notables entre la temporada seca de 2015 y las de los 2016 y 2017 (p= 0,048); exhibiendo una abundancia significativamente mayor durante la temporada seca 2015.
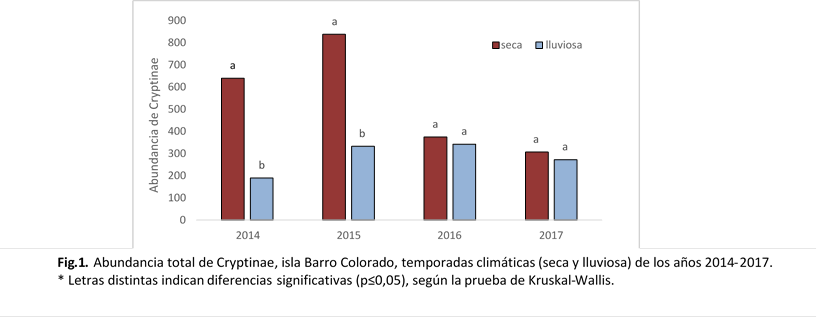 |
TABLA 2
Abundancia total de criptinos, Isla Barro Colorado. Temporadas seca y lluviosa
|
Año |
Abundancia por temporada |
|
|
Temporada seca |
Temporada lluviosa |
|
|
2014 |
640 abc |
190 a |
|
2015 |
838 b |
333 a |
|
2016 |
375 ac |
343 a |
|
2017 |
307 ac |
273 a |
|
Total |
2160 A |
1139 B |
Nota: Letras distintas indican diferencias significativas (p≤ 0,05), según la prueba de Kruskal-Wallis. Las letras minúsculas se utilizan para comparaciones estadísticas entre columnas, letras mayúsculas se utilizan para comparaciones estadísticas entre filas de dos columnas.
La riqueza de Cryptinae estuvo conformada por 15 morfoespecies (m. sp.) (Figs. 3a y 3b); el año 2014 presentó la mayor riqueza, con 14 m. sp., mientras que los años 2015, 2016 y 2017 mostraron la menor riqueza de Cryptinae (n = 13 m. sp.). En términos generales, se observó una mayor riqueza de Cryptinae durante la temporada seca (n = 14 m. sp.) en comparación con la temporada lluviosa (n = 13 M.E.). Las morfoespecies con mayor frecuencia de avistamientos fueron: Cryptinae sp.5 y sp.7, presentes en todos los años y épocas de recolección. En contraste las morfoespecies Cryptinae sp.13 y sp.15, estuvieron ausentes la mayor parte de la investigación (Fig. 2).
|
|
2014 |
2015 |
2016 |
2017 |
|
|||||||
|
|
Seca |
Lluv. |
Seca |
Lluv. |
Seca |
Lluv. |
Seca |
Lluv. |
TOTAL |
|||
|
Cryptinae sp.1 |
7 |
3 |
3 |
3 |
|
8 |
|
1 |
25 |
|||
|
Cryptinae sp.2 |
14 |
13 |
25 |
18 |
14 |
11 |
12 |
6 |
113 |
|||
|
Cryptinae sp.3 |
2 |
|
|
3 |
|
|
1 |
|
6 |
|||
|
Cryptinae sp.4 |
9 |
4 |
11 |
10 |
5 |
5 |
2 |
3 |
49 |
|||
|
Cryptinae sp.5 |
113 |
58 |
148 |
89 |
46 |
79 |
43 |
32 |
608 |
|||
|
Cryptinae sp.6 |
20 |
11 |
6 |
1 |
4 |
1 |
3 |
7 |
53 |
|||
|
264 |
30 |
392 |
84 |
166 |
95 |
111 |
122 |
1264 |
||||
|
Cryptinae sp.8 |
85 |
35 |
112 |
68 |
62 |
51 |
72 |
42 |
527 |
|||
|
Cryptinae sp.9 |
79 |
30 |
96 |
37 |
54 |
48 |
50 |
34 |
428 |
|||
|
Cryptinae sp.10 |
26 |
3 |
33 |
15 |
14 |
17 |
9 |
6 |
133 |
|||
|
Cryptinae sp.11 |
1 |
|
4 |
4 |
3 |
9 |
2 |
8 |
41 |
|||
|
Cryptinae sp.12 |
16 |
2 |
4 |
1 |
7 |
6 |
2 |
1 |
39 |
|||
|
1 |
1 |
|
|
|
1 |
|
|
3 |
||||
|
Cryptinae sp.14 |
3 |
|
|
|
|
2 |
|
1 |
6 |
|||
|
|
|
4 |
|
|
|
|
|
4 |
||||

Con respecto a los datos climáticos (Tabla 3), el año 2015 registró las temperaturas más altas, así como el menor registro de precipitaciones. En contraste, el año 2016 presentó la mayor precipitación entre todos los años analizados.
|
Año |
Temperatura (°C) |
Humedad relativa (%) |
Lluvia (mm) |
|
2014 |
25,89 |
91,5 |
2203 |
|
2015 |
26,26 |
92,2 |
1810 |
|
2016 |
25,75 |
91,7 |
2951 |
|
2017 |
25,74 |
93,3 |
2179 |
Nota: Para información adicional, consulte los Anexos 1, 2 y 3.
 |

DISCUSIÓN
A pesar de que la abundancia de criptinos registrada en esta investigación superó la reportada por Shapiro y Pickering en el año 2000, ambos estudios señalan a la subfamilia Cryptinae como la más abundante dentro de la familia Ichneumonidae en Barro Colorado. De manera similar, estudios realizados fuera de Panamá reportan resultados congruentes, destacando a Cryptinae por su alta abundancia en comparación con otras subfamilias (Veijalainen et al., 2013; Rodríguez et al., 2015; Álvarez et al., 2017; Pérez et al., 2018; Reátegui, 2019). No obstante, algunos estudios reportan resultados opuestos (Chay et al., 2006; Castillo et al., 2014; Orozco et al., 2019), incluyendo el estudio de Shapiro y Pickering (2000) en un bosque húmedo de Costa Rica. Estos resultados contrastantes podrían estar influenciados por diversos factores, como las condiciones climáticas (Rodríguez et al., 2015), gradientes altitudinales (Veijalainen et al., 2014) y la madurez del bosque (Shapiro y Pickering, 2000), en los cuales se llevaron a cabo las investigaciones.
Al igual que la mayoría de los insectos, los ichneumónidos son susceptibles a las variaciones ambientales de su entorno (Pérez et al., 2016); siendo los factores climáticos (precipitación y humedad) y la disponibilidad de hospederos, los principales elementos que regulan ocurrencia y proliferación (Shapiro y Pickering, 2000). Sin embargo, no todos los insectos reaccionan igual ante las mismas condiciones (Wolda,1978). Algunas especies pueden presentar una fuerte sincronía con los patrones climáticos, mientras que otras presentan fluctuaciones de abundancia que no están relacionadas con las estaciones climáticas (Wolda & Fisk, 1981). En el caso de la subfamilia Cryptinae, encontré una mayor preferencia por la temporada seca, en la cual presentó una abundancia significativamente mayor. Este resultado es opuesto al reportado por González et al., (2015) y a Shapiro y Pickering, 2000, quienes no reportaron diferencias significativas entre temporadas, pero si una ligera tendencia de la subfamilia por la temporada lluviosa.
En mi investigación encontré que la subfamilia Cryptinae se ve favorecida por condiciones de menor humedad y mayor temperatura, como las reportadas para el año 2015, influenciado en su totalidad por el fenómeno del niño (National Weather Service Climate Prediction Center. (n.d.)). Esto explicaría la mayor abundancia registrada en 2015 y las reducciones en años donde la precipitación fue superior al habitual, producto del efecto del fenómeno de la niña.
En relación con la riqueza, mi informe documenta 15 morfoespecies para Barro Colorado, una cifra inferior a la reportada por Castillo et al. (2014). Este resultado es relativamente bajo si se considera que la subfamilia es el grupo de Ichneumonidae más diverso del Neotrópico, con al menos 1,000 especies registradas (Hanson y Gauld, 2006). Esta discrepancia puede explicarse por el factor altitudinal, dado que la mayor riqueza de criptinos se registra entre los 1 033 y 1 709 metros sobre el nivel del mar (Veijalainen et al., 2014), mientras que la altitud máxima de Barro Colorado es de 137 metros sobre el lago Gatún (Pérez et al., 2005).
La notable riqueza de Cryptinae ha sido reiterada en diversas publicaciones mexicanas (Ruíz, 2010; Ruíz et al., 2011; Ruíz et al., 2014; Ruíz, 2015), las cuales destacan a esta subfamilia sobre otras por presentar el mayor número de géneros y especies descritas. Estos resultados son consistentes con los reportados por Orozco et al. (2019), quienes identificaron una mayor riqueza de géneros de Cryptinae en tres localidades de México. Asimismo, Castillo et al. (2019) señaló que el 30% del total de morfoespecies recolectadas en su estudio correspondía a dicha subfamilia.
En conclusión, la subfamilia Cryptinae se destaca como uno de los grupos más diversos y abundantes dentro de la familia Ichneumonidae, tanto en Barro Colorado como en otras regiones del Neotrópico. No obstante, su ocurrencia parece estar influenciada por factores climáticos, como la precipitación y temperatura, así como por gradientes altitudinales. Por lo tanto, es fundamental realizar estudios adicionales para comprender con mayor precisión los factores que regulan su riqueza y abundancia.
AGRADECIMIENTOS
Agradezco al Programa Regional de Becas del Servicio Alemán de Intercambio Académico (DAAD) por la beca completa; a Héctor Barrios, del Programa Centroamericano de Maestría en Entomología (PCMENT), y al Instituto Smithsonian de Investigaciones Tropicales (STRI), por el material biológico; a Paul Hanson por las claves taxonómicas, y a Samanta Orellana por su ayuda.
ÉTICA, CONFLICTO DE INTERESES Y DECLARACIÓN DE FINANCIAMIENTO
El autor declara que ha cumplido plenamente con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la producción del manuscrito; que no existen conflictos de interés de ningún tipo; que todas las fuentes financieras están completamente y claramente especificadas en la sección de agradecimientos; y que están de acuerdo con la versión final editada del artículo.
REFERENCIAS
Álvarez-Cabrera, G. M., Ruíz-Cancino, E., Coronado-Blanco, J. M., Treviño-Carreón, J., & Khalaim, A. I. (2017). Ichneumonidae (Hymenoptera) en diferentes tipos de vegetación del estado de Tamaulipas, México. Folia Entomológica Mexicana (nueva serie), 3(2), 42–53.
Álvarez-Cabrera, G. M., Ruíz-Cancino, E., Coronado-Blanco, J. M., Treviño-Carreón, J., & Khalaim, A. I. (2018). Propuesta de ichneumónidos (Hymenoptera) para el control biológico de insectos plaga en México. Agro Productividad, 10(9), 78–83. https://revista-agroproductividad.org/index.php/agroproductividad/article/view/979
Barone, J. (1998). Effects of light availability and rainfall on leaf production in a moist tropical forest in central Panama. Journal of Tropical Ecology, 14(3), 309–321.
Castillo-Flores, P. M., Ruíz-Cancino, E., Coronado-Blanco, J. M., Khalaim, A. I., & Myartseva, S. N. (2014). Ichneumonidae (Hymenoptera: Ichneumonoidea) en un bosque de Quercus de Tamaulipas, México. Dugesiana, 21(2), 161–174.
Castillo-Sánchez, L. E., Jiménez-Osornio, J. J., Delfín-González, H., Ramírez-Pech, J., Canul-Solís, J. R., González-Moreno, A., & Campos-Navarrete, M. J. (2019). Diversidad de Ichneumonoidea (Hymenoptera) en tres tipos de uso de suelo en un agroecosistema de producción múltiple en Xmatkuil, Yucatán, México. Revista Bio Ciencias, 6, e543. https://doi.org/10.15741/revbio.06.e543
Chay-Hernández, D. A., Delfín-González, H., & Parra-Tabla, V. (2006). Diversidad de comunidades de Ichneumonoidea (Hymenoptera) en un ambiente agrícola en el estado de Yucatán, México. Entomología Ambiental, 35(5), 1286–1297.
Çoruh, S., & Kolarov, J. (2022). A contribution to the Ichneumonidae (Hymenoptera) of Trabzon, III. Cryptinae [Trabzon’dan Ichneumonidae (Hymenoptera) Türlerine Katkılar III. Cryptinae]. Atatürk Üniversitesi Ziraat Fakültesi Dergisi, 53(1), 8–13. https://doi.org/10.54614/AUAF.2022.909906
Eisenmann, E. (1952). Annotated list of birds of Barro Colorado Island, Panama Canal Zone. Smithsonian Miscellaneous Collections, 117(5), 1–12.
Fincke, O. M., Higgins, L., & Rojas, E. (1990). Parasitism of Nephila clavipes (Araneae, Tetragnathidae) by an ichneumonid (Hymenoptera, Polysphinctini) in Panama. The Journal of Arachnology, 18(3), 321–329. http://www.jstor.org/stable/3705438
González-Moreno, A., Bordera, S., & Delfín-González, H. (2015). Spatio-temporal diversity of Cryptinae (Hymenoptera, Ichneumonidae) assemblages in a protected area of southeast Mexico. Journal of Insect Conservation, 19(6), 1153–1161. https://doi.org/10.1007/s10841-015-9830-1
Hanson, P., & Gauld, I. (2006). Hymenoptera de la región Neotropical. Memoirs of the American Entomological Institute, 77, 1–994.
Miranda, R., Santos-Murgas, A., Quintero-A, D., & Abrego-L., J. C. (2020). Insectos asociados a ovisacos de Argiope argentata (Fabricius, 1775) (Arachnida: Araneae) en Panamá. Intropica, 15(1), 8–15. https://doi.org/10.21676/23897864.3280
Mora-Prendas, J. (2025). Variación estacional de parasitoides idiobiontes y koinobiontes (Hymenoptera: Ichneumonidae) en la Isla Barro Colorado, Panamá. Cuadernos de Investigación UNED, 17(1). https://doi.org/10.22458/urj.v17i1.5648
Murgas, A. S., Lanuza, A. E., Zúñiga, R., & López, O. G. (2015). Primer registro para Panamá de Digonocryptus rufigaster (Szépligeti) (Hymenoptera: Ichneumonidae: Cryptinae), y sus hospederos, Anisopodus dispar Bates y Oreodera costaricensis Thomson (Coleoptera: Cerambycidae: Lamiinae). Revista Científica CENTROS (Panamá), 4(2), 31–39.
Murgas, A. S., Lanuza-Garay, A., Gutiérrez Lanzas, J., Flores, R., & Ábrego L., J. (2021). Cryptanura sp. (Hymenoptera: Ichneumonidae: Cryptinae), parasitoids of Taeniotes scalatus Gmelin (Coleoptera: Cerambycidae: Lamiinae) in Panamá. Aporte Santiaguino, 14(1), 43–51. https://doi.org/10.32911/as.2021.v14.n1.747
National Weather Service Climate Prediction Center. NWS CPC. (n.d.). Historical ONI values. Retrieved January 18, 2025, from https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php
Orozco-Peón, O., González-Moreno, A., Ruíz-Sánchez, E., & Tun-Suárez, J. M. (2019). Comunidades y gremios de parasitoides (Hymenoptera: Ichneumonidae) en cultivo de maíz y selva baja caducifolia circundante. Ecosistemas y Recursos Agropecuarios, 6(17), 195–205. https://doi.org/10.19136/era.a6n17.1977
Pádua, D. G., Moreira-Muñoz, A., Morales-Fierro, V., & Araujo, R. O. (2024). Chilean Darwin wasps (Ichneumonidae): Biogeographic relationships and distribution patterns. Insects, 15(6), 415. https://doi.org/10.3390/insects15060415
Pérez, R., Aguilar, S., Condit, R., Foster, R., Hubbell, S., & Lao, S. (2005). Metodología empleada en los censos de la parcela de 50 hectáreas de la Isla de Barro Colorado, Panamá (pp. 1–24). Centro de Ciencias Forestales del Trópico (CTFS) e Instituto Smithsonian de Investigaciones Tropicales (STRI). http://ctfs.si.edu/webatlas/onlinepubs/ParcelaBCIMetodo2005.pdf
Pérez-Urbina, B. A., Coronado-Blanco, J. M., Ruíz-Cancino, E., Venegas-Barrera, C. S., Correa-Sandoval, A., & Horta-Vega, J. V. (2018). Estructura comunitaria de Ichneumonidae (Hymenoptera) en un área de manglares de la zona costera de Tamaulipas, México. Revista Mexicana de Biodiversidad, 89(3), 823–835. https://doi.org/10.22201/ib.20078706e.2018.3.2437
R Core Team. (2017). R: A language and environment for statistical computing. The R Foundation for Statistical Computing. https://www.R-project.org
Reátegui Sinti, J. (2019). Eficiencia del uso de tres tipos de trampas entomológicas para la captura de parasitoides de la familia Ichneumonidae (Insecta: Hymenoptera) en la Reserva Nacional Allpahuayo Mishana Iquitos–Perú, Loreto–2014 [Tesis de licenciatura, Universidad Nacional de la Amazonía Peruana].
Rodríguez-Mota, A. J., Ruíz-Cancino, E., Ivanovich-Khalaim, A., Coronado-Blanco, J. M., & Treviño-Carreón, J. (2015). Diversidad de Ichneumonidae (Hymenoptera) en un bosque de Pinus spp. y Juniperus flaccida en Jaumave, Tamaulipas, México. Revista Mexicana de Biodiversidad, 86(4), 972–980. https://doi.org/10.1016/j.rmb.2015.08.003
Ruíz-Cancino, E., & Kasparyan, D. R. (2000). Una nueva especie del género Xorides Latreille (Hymenoptera: Ichneumonidae) del estado de Tamaulipas, México. Acta Zoológica Mexicana (nueva serie), 80, 233–239. https://www.scielo.org.mx/scielo.php?pid=S0065-17372000000200014&script=sci_arttext
Ruíz-Cancino, E., Coronado-Blanco, J. M., Khalaim, A. I., Toledo-Hernández, V. H., Flores-Palacios, A., Tovar-Sánchez, E., & García-Jiménez, J. (2012). Ichneumonidae (Hymenoptera) en el control natural y biológico de insectos en México. En E. Ruíz-Cancino & J. M. Coronado-Blanco (Eds.), Recursos naturales (pp. 244–252). Universidad Autónoma de Tamaulipas.
Ruíz-Cancino, E. (2015). La familia Ichneumonidae (Hymenoptera) en México. Entomología Mexicana, 2, 1–13.
Ruíz-Cancino, E., Coronado-Blanco, J. M., Rafaelevich-Kasparyan, D., & Ivanovich-Khalaim, A. (2011). La diversidad de avispas parasitoides en Tamaulipas: Familia Ichneumonidae. CienciaUAT, 6(1), 14–17.
Ruíz-Cancino, E., Coronado-Blanco, J. M., Kasparyan, D., & Khalaim, A. (2010). Ichneumonidae (Hymenoptera) en México. II Taller Internacional de Recursos Naturales, 1, 51–56.
Ruíz-Cancino, E., Rafaelevich-Kasparyan, D., González-Moreno, A., Khalaim, A. I., & Coronado-Blanco, J. M. (2014). Biodiversidad de Ichneumonidae (Hymenoptera) en México. Revista Mexicana de Biodiversidad, 85, 385–391. https://doi.org/10.7550/rmb.32448
Santos, B. F., & Hoppe, J. P. M. (2018). Filling gaps in species distributions through the study of biological collections: 415 new distribution records for Neotropical Cryptinae (Hymenoptera, Ichneumonidae). Revista Brasileira de Entomologia, 62(4), 288–291. https://doi.org/10.1016/j.rbe.2018.09.001
Shapiro, B. A., & Pickering, J. (2000). Rainfall and parasitic wasp (Hymenoptera: Ichneumonoidea) activity in successional forest stages at Barro Colorado Nature Monument, Panama, and La Selva Biological Station, Costa Rica. Agricultural and Forest Entomology, 2(1), 39–47. https://doi.org/10.1046/j.1461-9563.2000.00048.x
Veijalainen, A., Sääksjärvi, I. E., Erwin, T. L., Gómez, I. C., & Longino, J. T. (2013). Subfamily composition of Ichneumonidae (Hymenoptera) from western Amazonia: Insights into diversity of tropical parasitoid wasps. Insect Conservation and Diversity, 6(1), 28–37. https://doi.org/10.1111/j.1752-4598.2012.00185.x
Wolda, H., & Fisk, F. W. (1981). Seasonality of tropical insects. II Blataria in Panama. Journal of Animal Ecology, 50, 827–838.
Yu, D. S. K. (2005). Taxapad Ichneumonoidea. In Catalogue of Life (version May 2009). Consultado el 01 de mayo de 2025, en https://doi.org/10.48580/dgjc7-39v
 |
lang=ES-CR>
 |
|||
 |
|||