
Emisión de brotes y relación con clima en pitahaya (Selenicereus costaricensis; Cactaceae)
José Eladio
Monge-Pérez1![]() &
Michelle Loría-Coto2
&
Michelle Loría-Coto2![]()
1. Universidad de Costa Rica, Finca Experimental Interdisciplinaria de Modelos Agroecológicos, Turrialba, Cartago, Costa Rica; jose.mongeperez@ucr.ac.cr
2. Universidad Estatal a Distancia, Escuela de Ciencias Exactas y Naturales, carretera a Sabanilla, Mercedes de Montes de Oca, San José, Costa Rica; michelle_loria@yahoo.com
Recibido 20-II-2024 ● Corregido 30-IV-2024 ● Aceptado 19-V-2024
https://doi.org/10.22458/urj.v16i1.5190
|
ABSTRACT. “Sprout emission and relationship with climate in pitahaya (Selenicereus costaricensis; Cactaceae)”. Introduction: For the production of fruit crops, it is important to know the relationship between climatic variables and crop phenology. There are few studies on this topic in pitahaya cultivation in Costa Rica. Objective: to evaluate the type of relationship between climate and the emission of sprouts in the cultivation of pitahaya cv. San Ignacio. Methods: We carried out the research between 2017 and 2019. We evaluated the following variables: number of vegetative sprouts (NVS), number of reproductive sprouts (NRS), average temperature (Ta), minimum temperature (Tmin), maximum temperature (Tmax), relative humidity (RH), and rainfall (R). We calculated Spearman correlations between variables. Results: We did not find statistically significant correlations between NVS and the other analyzed variables. On the other hand, we observed a significant positive correlation between the NRS and minimum temperature (r=0.52**), rainfall (r=0.36*), and relative humidity (r=0.33*). These results indicate that as precipitation, relative humidity, and minimum temperature increase, the number of reproductive sprouts also increases. We additionally identified correlations between NVS, NRS, and climatic variables during the months prior to sprouting. Conclusion: No defined pattern was observed in the emission of vegetative sprouts, while the emission of reproductive sprouts occurred mainly during the initial and middle phases of the rainy season. Furthermore, it was observed that generally, during the emission of reproductive sprouts, few or no vegetative sprouts were produced.
Keywords: Central America, bloom, rainfall, drought, phenology.
|
RESUMEN. Introducción: Para la producción de frutales, es importante conocer la relación entre las variables climáticas y la fenología del cultivo. Hay pocos estudios sobre este tema en el cultivo de pitahaya en Costa Rica. Objetivo: Evaluar el tipo de relación entre la emisión de brotes en el cultivo de pitahaya cv. San Ignacio y el clima. Métodos: Realizamos la investigación entre los años 2017 al 2019. Evaluamos las siguientes variables: número de brotes vegetativos (NBV), número de brotes reproductivos (NBR), temperatura promedio (Tp), mínima (Tmín) y máxima (Tmáx); humedad relativa (HR) y precipitación pluvial (P). Calculamos las correlaciones de Spearman entre variables. Resultados: No encontramos correlaciones estadísticamente significativas entre el NBV y las demás variables analizadas. Por otro lado, observamos una correlación positiva significativa entre el NBR y las variables de temperatura mínima (r=0,52**), precipitación (r=0,36*) y humedad relativa (r=0,33*). Estos resultados indican que a medida que aumentan la precipitación, la humedad relativa y la temperatura mínima, también aumenta la cantidad de brotes reproductivos. Identificamos adicionalmente correlaciones entre NBV, NBR y las variables climáticas durante los meses previos a la brotación. Conclusión: No se observó un patrón definido en la emisión de brotes vegetativos, mientras que la emisión de brotes reproductivos ocurrió principalmente durante las fases inicial y media de la temporada de lluvias. Además, se observó que generalmente, durante la emisión de brotes reproductivos, se producían pocos o ningún brote vegetativo.
Palabras clave: Centroamérica, floración, lluvia, sequía, fenología.
|
La pitahaya [Selenicereus costaricensis (F.A.C. Weber) S. Arias y N. Korotkova ex Hammel, 2020] es una especie de la familia de las cactáceas, y su origen corresponde a las zonas subtropicales y tropicales de Centroamérica, México y Suramérica (Ángel et al., 2012; Montesinos et al., 2015). Esta planta se usa como alimento, especialmente por el fruto, pero también se utilizan las flores, semillas y brotes tiernos (Ángel et al., 2012; Montesinos et al., 2015). Con los frutos de esta especie se elaboran refrescos, jaleas, helados, gelatinas, mermeladas, cocteles, dulces, yogurt, entre otros (Huachi et al., 2015). Los frutos son ricos en fibras, flavonoides y vitaminas A y B (B1, B2 y B3), además de ser una fuente de potasio, fósforo y calcio (Ortiz & Takahashi, 2020).
En Centroamérica e Israel se cultivan principalmente las especies H. costaricensis, H. undatus, H. purpusii, H. polyrhizus y H. triangularis, las que son conocidas como pitahaya roja, cuyos frutos no presentan espinas (Ángel et al., 2012; Mizrahi, 2014; Montesinos et al., 2015). Entre las variedades de pitahaya que se cultivan en Nicaragua y Costa Rica están: “Orejona”, “Rosa”, “San Ignacio”, “Cebra”, “Nacional”, “Lisa” y “Crespa”, las cuales son de pulpa y cáscara roja (García & Quirós, 2010; López & Miranda, 2002; Mizrahi, 2014).
La pitahaya se adapta a ambientes áridos y secos, pues presenta mecanismos que le permiten captar agua y reducir la evapotranspiración (Ángel et al., 2012; Montesinos et al., 2015).
La fructificación de la pitahaya va de mayo-junio a octubre-noviembre en México y Nicaragua (Ángel et al., 2012; Cálix et al., 2014; Le Bellec et al., 2006), y en Costa Rica va de mayo a septiembre, aunque la producción se concentra entre junio y agosto (García & Quirós, 2010).
En esta planta, las yemas florales se producen luego del inicio de las lluvias (Ángel et al., 2012; Le Bellec et al., 2006), y emergen sobre todo a partir de los tallos que se desarrollaron anteriormente (Ángel et al., 2012).
La humedad relativa, temperatura promedio y precipitación pluvial, son variables del clima que generalmente afectan la fenología de las plantas, pues provocan el inicio de diversos mecanismos fisiológicos y la alteración del metabolismo (Chagas et al., 2019). El conocimiento sobre la fenología de una especie es muy importante con el fin de definir estrategias para su fitomejoramiento, conservación, y manejo agronómico (Chagas et al., 2019; Kishore, 2016).
La correlación de Spearman es una herramienta de análisis estadístico no paramétrico, que mide el grado de asociación que hay entre dos variables cuantitativas; por lo tanto, este procedimiento se puede usar cuando los datos presentan una distribución que no se ajusta al supuesto de normalidad (Di Rienzo et al., 2008; Mondragón, 2014). Varios autores han usado la correlación de Spearman para cuantificar la relación que existe entre el clima y la fenología de la palma aceitera (Chagas et al., 2019), y de la pitahaya (Marques et al., 2010; Monge-Pérez & Loría-Coto, 2022a; 2022b; 2022c).
En este ensayo evaluamos la relación entre las variables de clima y la emisión de brotes en pitahaya cv. San Ignacio, en San Mateo, Alajuela, Costa Rica.
MATERIALES Y MÉTODOS
Establecimos el ensayo en una finca de pitahaya (Selenicereus costaricensis; número de catálogo 231364, Herbario Nacional de Costa Rica), cv. “San Ignacio”, cultivada de manera orgánica, ubicada en San Mateo, Alajuela, Costa Rica, a una altitud de 232 msnm, que había sido sembrada el 1° de junio de 2016, con plantas propagadas por esqueje, sembradas a una distancia de 3m x 3m, y con postes vivos de jiñocuabe (Bursera simaruba) como tutores para las plantas. Fertilizamos las plantas con bocashi, a razón de 1,5kg/planta/año; aplicamos 500g de bocashi por planta en tres épocas de cada año (junio, agosto y octubre).
Hicimos un análisis de suelo a nivel general de la parcela, en el Laboratorio de Suelos, Centro de Investigaciones Agronómicas, Universidad de Costa Rica. Evaluamos diversas características a nivel físico, químico y microbiológico. Tomamos 20 submuestras, a una profundidad de 0-20cm, para conformar la muestra.
Seleccionamos aleatoriamente 10 plantas de pitahaya en la parcela para realizar las observaciones. Las visitas se llevaron a cabo cada dos semanas, durante las cuales evaluamos el número de brotes vegetativos (NBV) y brotes reproductivos (NBR) nuevos por planta. Empleamos la metodología de evaluación descrita previamente (Monge-Pérez & Loría-Coto, 2022a). Realizamos las evaluaciones desde el 8 de febrero de 2017 hasta el 26 de diciembre de 2019.
Además, solicitamos al Instituto Meteorológico Nacional (IMN) los datos diarios en San Mateo para el periodo 2017-2019, de las siguientes variables de clima: Temperatura mínima (Tmín, en °C); Temperatura máxima (Tmáx, en °C); Temperatura promedio (Tp, en °C); Precipitación pluvial (P, en mm); y Humedad relativa (HR, en %).
Para cada variable calculamos el promedio mensual, y ejecutamos la prueba de Shapiro-Wilks modificado a todos los datos, y confirmamos que ninguna variable se ajustó al supuesto de normalidad, por lo que utilizamos la correlación de Spearman para analizar la relación existente entre variables, según la descripción de Chagas et al. (2019), mediante el uso del software Infostat versión 2008 (Di Rienzo et al., 2008); destacamos aquellas correlaciones que tuvieron significancia estadística.
Además, para NBV y NBR, calculamos las correlaciones de Spearman en relación con las demás variables, según el registro de datos de los doce meses anteriores a la brotación, de acuerdo con la metodología descrita en un trabajo anterior (Monge-Pérez & Loría-Coto, 2022a).
RESULTADOS
El suelo es arcilloso (con 55% de arcillas), con una alta capacidad efectiva de intercambio catiónico (13,49cmol(+)/L), y cuya limitante más importante fue un contenido de fósforo bajo (4mg/L) (Tabla 1).
Con respecto a los resultados de las diversas variables de clima, la precipitación (P) osciló entre 0-168mm, y su promedio fue de 6,16mm (Fig. 1); la P anual fue más alta durante el 2017, y fue más baja en el 2018, mientras que en el año 2019 mostró un valor intermedio entre ambos años precedentes (Fig. 2). La HR se estableció entre 37-100%, y su promedio fue de 82,43% (Fig. 3); la Tp varió entre 21,8-30,0°C, con un promedio de 25,59°C; la Tmáx osciló entre 24,0-39,4°C, y su promedio fue de 33,21°C; y la Tmín varió entre 15,2-26,4°C, con un promedio de 20,39°C (Fig. 4).
En cuanto a la emisión de brotes vegetativos (BV) (Fig. 5), la máxima producción de brotes/planta se registró en julio (1,17), septiembre (1,00), octubre (1,00) y noviembre (1,00) del año 2017; durante marzo (1,33) y noviembre (0,80) del año 2018; y durante enero (0,80), abril (1,30), mayo (1,00), septiembre (1,40), noviembre (1,40) y diciembre (1,20) del año 2019.
TABLA 1.
Características del suelo, San Mateo
|
Características |
Valor obtenido |
Valor de referencia |
|
Físicas |
||
|
Arcilla (%) |
55 |
nd |
|
Limo (%) |
23 |
nd |
|
Arena (%) |
22 |
nd |
|
Clase textural |
Arcilloso |
nd |
|
Químicas |
||
|
pH (H2O) |
5,9 |
5,5 |
|
Acidez (cmol(+)/L) |
0,12 |
0,5 |
|
Ca (cmol(+)/L) |
10,37 |
4 |
|
Mg (cmol(+)/L) |
2,14 |
1 |
|
K (cmol(+)/L) |
0,86 |
0,2 |
|
CICE (cmol(+)/L) |
13,49 |
5 |
|
SA (%) |
0,9 |
nd |
|
P (mg/L) |
4 |
10 |
|
Zn (mg/L) |
6,9 |
3 |
|
Cu (mg/L) |
20 |
1 |
|
Fe (mg/L) |
91 |
10 |
|
Mn (mg/L) |
27 |
5 |
|
CE (mS/cm) |
0,3 |
1,5 |
|
C (%) |
4,65 |
nd |
|
N (%) |
0,44 |
nd |
|
Relación C/N |
10,6 |
nd |
|
S (mg/L) |
17 |
12 |
|
MO (%) |
6,65 |
nd |
|
Microbiológicas |
||
|
Hongos (UFC/g) |
9,9 x 104 |
nd |
|
Hongos dominantes |
Fusarium sp., Aspergillus sp., Trichoderma sp., Penicillium sp. |
nd |
Nota: CICE: capacidad efectiva de intercambio catiónico = Acidez+Ca+Mg+K; SA: saturación de la acidez = (Acidez/CICE)*100; CE: conductividad eléctrica; MO: materia orgánica; nd: no dato; valor de referencia: valor que se considera como deseable en el suelo, según Centro de Investigaciones Agronómicas, Universidad de Costa Rica.
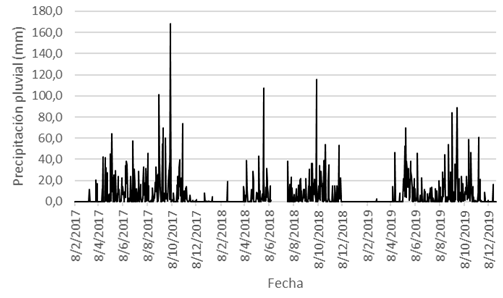
Fig. 1. Precipitación pluvial (P, en mm), San Mateo.
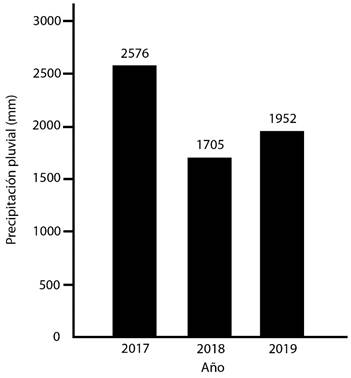
Fig. 2. Precipitación pluvial anual (en mm), San Mateo (2017-2019).
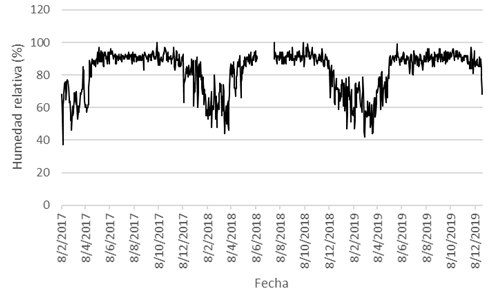
Fig. 3. Humedad relativa (HR, en %), San Mateo
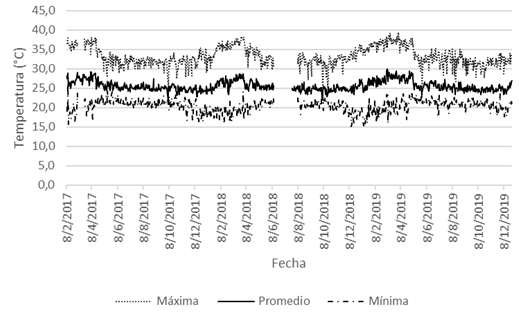
Fig. 4. Temperatura (promedio, máxima y mínima; °C), San Mateo
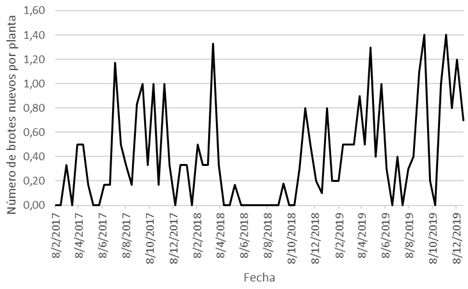
Fig. 5. Evolución de la emisión de BV nuevos en pitahaya cv. San Ignacio.
En relación con la emisión de brotes reproductivos (BR) (Fig. 6), la máxima producción de brotes/planta se registró durante abril (0,33), junio (2,00) y julio (0,82) del año 2018; y durante abril (0,30), mayo (3,80), julio (1,50) y septiembre (1,00) del año 2019.

Fig. 6. Evolución de la emisión de BR nuevos en pitahaya cv. San Ignacio.
Con respecto a la relación entre P y la emisión de BR, en el año 2018 observamos varios episodios de producción de BR (Fig. 7), y el primero de ellos ocurrió 13 días después de la primera lluvia de la temporada, que sucedió el 5 de abril. A pesar de que el 24 de febrero hubo un aguacero importante, fue una precipitación aislada, pues luego se presentaron 39 días sin lluvias, hasta el 5 de abril. El punto de máxima emisión de brotes reproductivos se alcanzó el 1° de junio, aproximadamente una semana después de un fuerte aguacero.
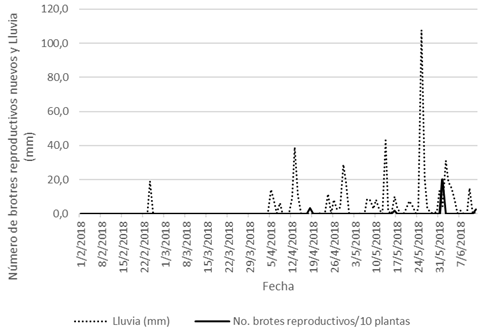
Fig. 7. Relación entre P (mm) y la emisión de BR nuevos (1° de febrero de 2018 a 12 de junio de 2018).
En cuanto a la relación entre P y la emisión de BR, en el año 2019, el 3 de marzo ocurrió una leve lluvia, y la siguiente precipitación fue el 13 de abril, y los primeros BR se emitieron tres días después (Fig. 8). Entre el 8 y el 13 de mayo se acumuló bastante lluvia (110,9mm), y el día 14 de mayo se obtuvo el punto de máxima emisión de brotes reproductivos, con un promedio de 3,80 BR por planta.
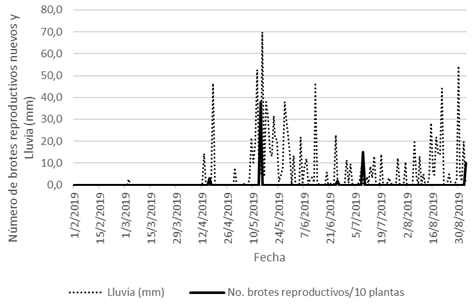
Fig. 8. Relación entre P (mm) y la emisión de BR nuevos (1° de febrero de 2019 al 3 de septiembre de 2019).
Los datos de la correlación entre la emisión de brotes y las variables de clima se muestran en la Tabla 2; no hallamos correlaciones significativas entre NBV y las demás variables.
Encontramos correlaciones positivas entre el NBR y las variables Tmín (r=0,52**), P (r=0,36*), y HR (r=0,33*). Además, hallamos correlaciones con significancia estadística entre las variables de clima, excepto entre Tmín y Tp.
TABLA 2
Correlación de Spearman entre las variables de clima y la emisión de brotes
|
|
NBV |
NBR |
Tp |
Tmáx |
Tmín |
P |
HR |
|
NBV |
1 |
-0,01 ns |
0,10 ns |
-0,05 ns |
-0,01 ns |
0,05 ns |
-0,13 ns |
|
NBR |
|
1 |
0,23 ns |
0,12 ns |
0,52** |
0,36* |
0,33* |
|
Tp |
|
|
1 |
0,79** |
-0,11 ns |
-0,42** |
-0,70** |
|
Tmáx |
|
|
|
1 |
-0,59** |
-0,77** |
-0,94** |
|
Tmín |
|
|
|
|
1 |
0,84** |
0,72** |
|
P |
|
|
|
|
|
1 |
0,83** |
|
HR |
|
|
|
|
|
|
1 |
Nota: Los datos corresponden a promedios mensuales. NBV: número de brotes vegetativos; NBR: número de brotes reproductivos; Tmín: temperatura mínima (°C); Tmáx: temperatura máxima (°C); Tp: temperatura promedio (°C); HR: humedad relativa (%); P: precipitación pluvial (mm). *: p≤0,05; **: p≤0,01; ns: no significativa.
Los datos de la correlación entre la emisión de BV y las otras variables se muestran en la Tabla 3, según los meses que anteceden a los brotes vegetativos (MABV). Encontramos una correlación positiva entre NBV y NBR a los 7 y 9 MABV.
También, NBV obtuvo correlación positiva con Tp a 8 MABV, y también con Tmáx entre 8 y 11 MABV. Además, hallamos una correlación de signo positivo entre Tmín y NBV entre 4 y 6 MABV, y de signo negativo entre ellas a 11-12 MABV.
TABLA 3
Correlación entre el NBV y las otras variables, en los meses anteriores a la emisión de BV
|
Primera variable |
Momento evaluación segunda variable (MABV) |
Segunda variable |
|||||
|
NBR |
Tp |
Tmáx |
Tmín |
P |
HR |
||
|
NBV |
1 |
ns |
ns |
ns |
ns |
ns |
ns |
|
2 |
ns |
ns |
ns |
ns |
ns |
ns |
|
|
3 |
ns |
ns |
ns |
ns |
ns |
ns |
|
|
4 |
ns |
ns |
ns |
0,37* |
ns |
ns |
|
|
5 |
ns |
ns |
ns |
0,47** |
ns |
ns |
|
|
6 |
ns |
ns |
ns |
0,37* |
ns |
ns |
|
|
7 |
0,44* |
ns |
ns |
ns |
ns |
ns |
|
|
8 |
ns |
0,42* |
0,45* |
ns |
ns |
ns |
|
|
9 |
0,44* |
ns |
0,42* |
ns |
ns |
ns |
|
|
10 |
ns |
ns |
0,46* |
ns |
ns |
ns |
|
|
11 |
ns |
ns |
0,42* |
-0,42* |
ns |
ns |
|
|
12 |
ns |
ns |
ns |
-0,57** |
ns |
ns |
|
Nota: Los datos corresponden a promedios mensuales. MABV: meses que anteceden a los brotes vegetativos; NBV: número de brotes vegetativos; NBR: número de brotes reproductivos; Tmín: temperatura mínima (°C); Tmáx: temperatura máxima (°C); Tp: temperatura promedio (°C); HR: humedad relativa (%); P: precipitación pluvial (mm). *: p≤0,05; **: p≤0,01; ns: no significativa.
Los datos de la correlación entre la emisión de BR y las otras variables se muestran en la Tabla 4, según los meses que anteceden a los brotes reproductivos (MABR). Obtuvimos una correlación positiva entre NBR y NBV a los 5 MABR.
TABLA 4
Correlación entre el NBR y las otras variables, en los meses anteriores a la emisión de BR
|
Primera variable |
Momento evaluación segunda variable (MABR) |
Segunda variable |
|||||
|
NBV |
Tp |
Tmáx |
Tmín |
P |
HR |
||
|
NBR |
1 |
ns |
0,50** |
0,38* |
0,42** |
ns |
ns |
|
2 |
ns |
0,64** |
0,54** |
ns |
ns |
ns |
|
|
3 |
ns |
0,69** |
0,68** |
ns |
ns |
-0,42* |
|
|
4 |
ns |
0,46** |
0,69** |
-0,38* |
-0,35* |
-0,48** |
|
|
5 |
0,36* |
ns |
0,47** |
-0,54** |
-0,43* |
-0,36* |
|
|
6 |
ns |
ns |
ns |
ns |
ns |
ns |
|
|
7 |
ns |
-0,41* |
ns |
ns |
ns |
ns |
|
|
8 |
ns |
-0,37* |
ns |
ns |
0,44* |
0,50** |
|
|
9 |
ns |
-0,45* |
-0,42* |
0,42* |
0,61** |
0,67** |
|
|
10 |
ns |
ns |
ns |
0,62** |
0,61** |
0,71** |
|
|
11 |
ns |
ns |
ns |
0,70** |
0,60** |
0,67** |
|
|
12 |
ns |
ns |
ns |
0,60** |
0,51* |
0,43* |
|
Nota: Los datos corresponden a promedios mensuales. MABR: meses que anteceden a los brotes reproductivos; NBV: número de brotes vegetativos; NBR: número de brotes reproductivos; Tmín: temperatura mínima (°C); Tmáx: temperatura máxima (°C); Tp: temperatura promedio (°C); HR: humedad relativa (%); P: precipitación pluvial (mm). *: p≤0,05; **: p≤0,01; ns: no significativa.
Encontramos una correlación positiva entre la temperatura promedio (Tp) y el número de brotes reproductivos (NBR) entre 1 y 4 MABR, y una correlación negativa entre ellas entre 7 y 9 MABR. Además, encontramos una correlación de signo positivo entre Tmáx y NBR entre 1 y 5 MABR, y de signo negativo entre ellas a 9 MABR.
Obtuvimos una correlación positiva entre Tmín y NBR a 1 MABR, así como entre 9 y 12 MABR, pero también una correlación negativa entre ellas a 4-5 MABR. Asimismo, encontramos una correlación de signo positivo entre NBR y P entre 8 y 12 MABR, y una correlación de signo negativo entre dichas variables a 4 y 5 MABR. Además, hallamos una correlación negativa entre HR y NBR entre 3 y 5 MABR, así como una correlación positiva entre ellas entre 8 y 12 MABR.
DISCUSIÓN
Expertos en investigación agronómica, específicamente enfocados en el cultivo de pitahaya, han señalado que, en esta especie, el punto óptimo de fijación neta de CO2 se produce cuando la temperatura diurna es de 30°C y la nocturna es de 20°C (Mizrahi, 2014; Nobel & De la Barrera, 2002a); durante la ejecución de nuestro ensayo la Tp y la Tmín promedio se ubicaron en ese rango, pero la Tmáx promedio estuvo por encima de dicho rango.
Es evidente que la P causó un incremento en la HR y en la Tmín, así como una reducción en la Tp y la Tmáx, lo que corresponde a consecuencias lógicas debido a la presencia de lluvias. Durante la temporada de lluvias, los valores de HR que hallamos fueron siempre superiores a 80%.
No hallamos un patrón predecible de emisión de BV, excepto que en el mes de noviembre de los tres años del estudio se verificó la producción de este tipo de brotes.
En otros ensayos se informó que la pitahaya produce anualmente varios ciclos de brotación vegetativa (Kishore, 2016; Trivellini et al., 2020); esto fue confirmado en nuestro ensayo.
En el año 2017 la emisión de BR fue muy baja, pues las plantas tenían solamente un año de edad. En el período evaluado, los BR se emitieron en la época de lluvias, pero únicamente en las fases inicial y media, pero nunca en la fase final de dicha temporada; en 2017 se emitieron el 14 de junio y el 9 de agosto, pero las lluvias continuaron hasta mediados de noviembre; en 2018 se emitieron entre el 18 de abril y el 9 de agosto, pero la temporada de lluvias concluyó en los primeros días de diciembre; y en 2019 se emitieron entre mediados de abril e inicios de septiembre, pero la temporada de lluvias concluyó al final del mes de noviembre.
Los resultados que encontramos con respecto a la producción de BR fueron similares a los hallados en otra investigación, en la cual la floración inició luego de que empezara la estación de lluvias (Marques et al., 2010), por lo que la mayor emisión de BR se dio cuando se presentaron los mayores valores de HR, pues la floración ocurrió solamente cuando la HR fue cercana al 80% (Marques et al., 2010); este fenómeno se observó también en la presente investigación, dado que durante el período de floración, la HR fue mayor a 80%.
El hecho de que no haya floración al final de la estación lluviosa se podría explicar porque las plantas dirigen sus fotoasimilados hacia los frutos que se produjeron a partir de los BR que se emitieron en las fases inicial y media de esa estación, por lo que la cantidad de fotoasimilados no es suficiente para la producción de BR en la etapa final de la misma.
En el año 2018 registramos tres ciclos de floración, y en el año 2019 cuatro ciclos. Este resultado es similar al reportado por otros investigadores, quienes hallaron 3-5 ciclos anuales de floración (Kishore, 2016; Marques et al., 2011; Martínez-Ruiz et al., 2017; Nerd et al., 2002), pero menor a lo obtenido por otros autores, entre 5-9 ciclos anuales (Ángel et al., 2012; Osuna-Enciso et al., 2016; Silva et al., 2015). En un ensayo en Taiwan, la emisión de BR osciló entre 3-6 ciclos anuales, dependiendo del genotipo (Ha et al., 2018), mientras que en otra investigación se halló que los clones de pitahaya mostraron entre 2 y 10 ciclos de floración al año (Mizrahi, 2014).
El desarrollo vegetativo compitió con el reproductivo, dado que, en los meses de mayor emisión de BR, la emisión de BV que encontramos fue baja o nula; las principales excepciones a este comportamiento se dieron en abril y septiembre de 2019, en que la emisión de ambos tipos de brotes fue similar en las mismas fechas. A pesar de que en algunas plantas hallamos BV y BR en el mismo momento, lo más frecuente fue que, cuando la planta presentaba BR, no mostraba BV; este mismo comportamiento fue reportado por otros autores (Trivellini et al., 2020). También encontramos que, al aumentar la edad de la plantación, se incrementó el NBR por planta.
La planta de pitahaya es isohídrica, por lo que la sequía provoca solamente una leve disminución en su contenido de agua, debido al efecto del ABA (ácido abscísico) que se produce a nivel radical, y que provoca que los estomas se cierren (Nobel & De la Barrera, 2002b). En esas condiciones, un pequeño evento de lluvia no es capaz de provocar una apertura de estomas suficientemente prolongada que ayude a disminuir la cantidad de agua contenida en la planta de manera significativa (Nobel & De la Barrera, 2002b); es probable que este fuera el motivo por el que no encontramos que el aguacero de febrero de 2018 haya causado la emisión de BR, sino que fue necesario que la temporada de lluvias iniciara, para que los estomas de las plantas se mantuvieran abiertos y con ello lograr la maximización de la fijación de carbono. La respuesta inmediata de la planta de pitahaya ante la incidencia de las lluvias, se explica por su capacidad de absorber más carbono luego de un evento de sequía (Nobel & De la Barrera, 2002b). La pitahaya, al poseer el mecanismo ácido de las crasuláceas (CAM), presenta durante la noche su apertura máxima de estomas y también su mayor fijación neta de carbono (Nobel & De la Barrera, 2002b); bajo condiciones de estrés, el CAM ayuda a conservar la fotosíntesis y aumentar la eficiencia del uso del agua, pues los estomas están cerrados en el día, y esto le concede a esta especie adaptación a ambientes de alta temperatura y baja humedad, así como tolerancia a la sequía (Trivellini et al., 2020). En otro ensayo se informó que la tasa máxima fotosintética en pitahaya sucede en la estación de lluvias (Ortiz-Hernández et al., 2012).
Otros autores informaron que, bajo condiciones secas, se reduce la conductividad hidráulica en la pitahaya, pero ella regresa a valores semejantes de condiciones de buena humedad luego de tres días de haberse logrado nuevamente el suministro hídrico (Nobel & De la Barrera, 2002b). En pitahaya, otros investigadores encontraron que la emisión de BR se dio dos meses después del abastecimiento de agua a las plantas, producto de riego o de la lluvia (Martínez-Ruiz et al., 2017).
Nuestro resultado de que NBV no se correlacionó en forma significativa con las demás variables, también fue obtenido en otros ensayos con otras variedades de pitahaya (Monge-Pérez & Loría-Coto, 2022a; 2022b; 2022c); esto confirma que no se encontró un patrón característico o definido en la emisión de BV, relacionado con una variable climática.
Encontramos en nuestro ensayo que, conforme aumenta la temperatura mínima, la precipitación y la humedad relativa, se produce un mayor número de BR nuevos; esto confirma que los BR se emiten únicamente en la temporada de lluvias, momento en que aumentó también la Tmín y la HR.
En relación con las correlaciones entre NBR y las variables climáticas, de manera contraria a lo que encontramos en este trabajo, en otro ensayo en pitahaya se informó que la variable Tp mostró una correlación positiva alta (r=0,75) con el NBR (Marques et al., 2010); la explicación de esto es que en el sitio en Brasil en que se realizó el experimento presenta las cuatro estaciones, y los BR aparecieron mayoritariamente durante el verano (Tp = 20-25°C), y de ninguna forma cuando la Tp fue inferior a 20°C.
Para la relación entre floración y T en pitahaya, se informó que una temperatura de 34-38°C redujo de manera considerable la emisión de BR (Nerd et al., 2002); en el presente trabajo, en la estación seca la Tmáx registró valores mayores a 35°C, pero la Tp no fue superior a 30°C en ningún momento. Por otra parte, se ha informado que, con una temperatura inferior a 15°C, la actividad fisiológica de la pitahaya se reduce (Martínez-Ruiz et al., 2017).
En otro trabajo en pitahaya, se informó que la emisión de BR sucedió luego del período del año en que se presentaron las mayores temperaturas (Martínez-Ruiz et al., 2017); esto también ocurrió en la presente investigación.
En otro trabajo se informó que, además de la Tp, las variables HR y P también tienen una influencia importante en la emisión de BR (Marques et al., 2010); en nuestra investigación se obtuvo el mismo resultado con respecto a P y HR. Varios autores informaron que la emisión de BR se dio al inicio de la estación de lluvias y cuando ocurrieron temperaturas altas (mayores a 20°C) (Osuna-Enciso et al., 2016; Silva et al., 2015), y también cuando aumentó la HR (Osuna-Enciso et al., 2016); esos resultados son similares a los hallados en nuestra investigación con respecto a P, Tmín y HR.
Otros autores informaron que la T que se produce antes de la emisión de BR interfiere con el inicio de la floración, pues los BR empezaron a emitirse 2 meses luego de que Tp fuese mayor a 20°C (Marques et al., 2010). Por otra parte, en nuestra investigación la Tp fue mayor a 20°C en todo momento, lo que es una posible explicación de la no existencia de correlación entre Tp y NBR.
En otro trabajo, la floración sucedió cuando se presentó P, HR alta (mayor a 80%), una T moderada (28°C), y días largos (Kishore, 2016); lo mismo hallamos en nuestro trabajo.
Con respecto a los BV, hallamos que un incremento en Tp y Tmáx afectó positivamente el NBV emitido luego de 8 meses. Asimismo, que un aumento en Tmín afectó positivamente el NBV producido 4-6 meses después, pero negativamente el NBV emitido luego de 11-12 meses. También hallamos que un mayor NBR afectó en forma positiva el NBV producido luego de 7 y 9 meses.
En relación con los BR, encontramos que un incremento en NBV afectó de manera positiva el NBR emitido luego de 5 meses. Asimismo, que un incremento en Tp y Tmáx afectó positivamente el NBR producido luego de 1-4 meses, pero negativamente sobre el NBR emitido luego de 9 meses. Además, que un aumento en Tmín provocó un incremento en el NBR producido luego de un mes, y luego de 9-12 meses, pero ocasionó una reducción en el NBR emitido luego de 4-5 meses.
Por otra parte, hallamos que un incremento en P provocó un mayor NBR luego de 8-12 meses, y una reducción en NBR luego de 4-5 meses; esto respalda el hecho de que no hubiera emisión de BR al final de la temporada lluviosa (luego del final de agosto o del inicio de septiembre), dado que el inicio de dicha temporada sucedió en abril de cada año. De la misma forma, encontramos que un incremento en HR afectó en forma negativa el NBR emitido luego de 3-5 meses, pero de manera positiva el NBR producido luego de 8-12 meses.
No hallamos correlaciones significativas entre NBV y P, ni entre NBV y HR, en los meses anteriores a la brotación vegetativa. Por el contrario, en otros trabajos sí se hallaron correlaciones significativas entre dichas variables para otras variedades de pitahaya (Monge-Pérez & Loría-Coto, 2022a; 2022b; 2022c).
Por lo tanto, podemos concluir que no encontramos correlaciones estadísticamente significativas entre el NBV y las demás variables analizadas.
Por otro lado, observamos una correlación positiva significativa entre el NBR y las variables de temperatura mínima (r=0,52**), precipitación (r=0,36*) y humedad relativa (r=0,33*). Estos resultados indican que a medida que aumentan la precipitación, la humedad relativa y la temperatura mínima, también aumenta la cantidad de brotes reproductivos. Identificamos adicionalmente correlaciones entre NBV, NBR y las variables climáticas durante los meses previos a la brotación.
No se observó un patrón definido en la emisión de brotes vegetativos, mientras que la emisión de brotes reproductivos ocurrió principalmente durante las fases inicial y media de la temporada de lluvias. Además, se observó que generalmente, durante la emisión de brotes reproductivos, se producían pocos o ningún brote vegetativo.
AGRADECIMIENTOS
Agradecemos a Patricia Oreamuno, Carlos Blanco y Mario Monge por su colaboración en la realización de este trabajo; al Instituto Meteorológico Nacional por la cesión de los datos meteorológicos; y a la Universidad de Costa Rica por la colaboración financiera.
ÉTICA, CONFLICTO DE INTERESES Y DECLARACIÓN DE FINANCIAMIENTO
Declaramos haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la preparación de este documento; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, estamos de acuerdo con la versión editada final de esta publicación. El respectivo documento legal firmado se encuentra en los archivos de la revista.
La declaración de contribución de cada autor es la siguiente: J.E.M.P.: Diseño del estudio, recolección y análisis de datos. M.L.C.: Análisis de datos. Todos los coautores.: preparación y aprobación final del manuscrito.
REFERENCIAS
Ángel, A. L., Estrada, C. A., Rebolledo, A., & Zetina, R. (2012). Pitahayas: patrimonio biocultural para diversificar la agricultura y la alimentación. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias.
Cálix, H., Castillo, R., & Caamal, H. J. (2014). Caracterización de la producción de pitahaya (Hylocereus spp.) en la zona maya de Quintana Roo, México. Agroecología, 9(1/2), 123-132.
Chagas, K. P., Carvalho, B. L., Guerra, C. A., Silva, R. A., & Vieira, F. A. (2019). Fenologia do dendezeiro e correlações com variáveis climáticas. Ciência Florestal, 29(4), 1701-1711.
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., González, L., Tablada, M., & Robledo, C. W. (2008). InfoStat, versión 2008. Córdoba, Argentina: Grupo Infostat, FCA, Universidad Nacional de Córdoba.
García, M. E., & Quirós, O. (2010). Análisis del comportamiento de mercado de la pitahaya (Hylocereus undatus) en Costa Rica. Tecnología en Marcha, 23(2), 14-24.
Ha, T. D., Oanh, L. T., & Yen, C. (2018). Flowering phenology and mating system of a red skin pitaya (Hylocereus spp.) germplasm collection in Taiwan. Asian Journal of Advances in Agricultural Research, 7(3), 1-8.
Huachi, L., Yugsi, E., Paredes, M. F., Coronel, D., Verdugo, K., & Coba, P. (2015). Desarrollo de la pitahaya (Cereus sp.) en Ecuador. La Granja: Revista de Ciencias de la Vida, 22(2), 50-58.
Kishore, K. (2016). Phenological growth stages of dragon fruit (Hylocereus undatus) according to the extended BBCH-scale. Scientia Horticulturae, 213, 294-302.
Le Bellec, F., Vaillant, F., & Imbert, E. (2006). Pitahaya (Hylocereus spp.): a new fruit crop, a market with a future. Fruits, 61(4), 237-250.
López, H., & Miranda, A. (2002). Cultivo de la pitahaya (Vol. Guía tecnológica 6). Instituto Nicaragüense de Tecnología Agropecuaria.
Marques, V. B., Moreira, R. A., Ramos, J. D., Araújo, N. A., & Silva, F. O. (2011). Fenologia reprodutiva de pitaia vermelha no município de Lavras, MG. Ciência Rural, 41(6), 984-987.
Marques, V. B., Ramos, J. D., Araújo, N. A., & Moreira, R. A. (2010). Correlação dos fatores ambientais e o período reprodutivo da pitaia (Hylocereus undatus) em Lavras-MG. XXI Congresso Brasileiro de Fruticultura, (pp. 1-5). Natal, RN, Brasil.
Martínez-Ruiz, E. R., Tijerina-Chávez, L., Becerril-Román, A. E., Rebolledo-Martínez, A., Velasco-Cruz, C., & Ángel-Pérez, A. L. (2017). Fenología y constante térmica de la pitahaya (Hylocereus undatus Haw. Britt. & Rose). Agro Productividad, 10(9), 3-8.
Mizrahi, Y. (2014). Vine-cacti pitayas - the new crops of the world. Revista Brasileira de Fruticultura, 36(1), 124-138.
Mondragón, M. A. (2014). Uso de la correlación de Spearman en un estudio de intervención en fisioterapia. Movimiento Científico, 8(1), 98-104.
Monge-Pérez, J. E., & Loría-Coto, M. (2022a). Pitahaya (Hylocereus sp.) cv. Amarilla: producción de brotes y relación con variables climáticas. Producción Agropecuaria y Desarrollo Sostenible, 11(1), 61-79.
Monge-Pérez, J. E., & Loría-Coto, M. (2022b). Producción de brotes en pitahaya (Hylocereus sp.) cultivada en Guanacaste, Costa Rica: relación con variables climáticas. Intersedes, 23(48), 94-114.
Monge-Pérez, J. E., & Loría-Coto, M. (2022c). Producción de brotes en pitahaya (Hylocereus sp.): correlación con variables climáticas. Avances en Investigación Agropecuaria, 26, 94-108.
Montesinos, J. A., Rodríguez-Larramendi, L., Ortiz-Pérez, R., Fonseca-Flores, M. A., Ruíz, G., & Guevara-Hernández, F. (2015). Pitahaya (Hylocereus spp.): un recurso fitogenético con historia y futuro para el trópico seco mexicano. Cultivos Tropicales, 36(especial), 67-76.
Nerd, A., Sitrit, Y., Kaushik, R. A., & Mizrahi, Y. (2002). High summer temperatures inhibit flowering in vine pitaya crops (Hylocereus spp.). Scientia Horticulturae, 96(1-4), 343-350.
Nobel, P. S., & De la Barrera, E. (2002a). High temperatures and net CO2 uptake, growth, and stem damage for the hemiepiphytic cactus Hylocereus undatus. Biotropica, 34(2), 225-231.
Nobel, P. S., & De la Barrera, E. (2002b). Stem water relations and net CO2 uptake for a hemiepiphytic cactus during short-term drought. Environmental and Experimental Botany, 48, 129-137.
Ortiz, T. A., & Takahashi, L. S. (2020). Pitaya fruit quality (Hylocereus undatus [Haworth] Britton & Rose) according to physiological maturity. A review. Revista Colombiana de Ciencias Hortícolas, 14(1), 63-75.
Ortiz-Hernández, Y. D., Livera-Muñoz, M., Carrillo-Salazar, J. A., Valencia-Botin, A. J., & Castillo-Martínez, R. (2012). Agronomical, physiological, and cultural contributions of pitahaya (Hylocereus spp.) in Mexico. Israel Journal of Plant Sciences, 60, 359-370.
Osuna-Enciso, T., Valdez-Torres, J. B., Sañudo-Barajas, J. A., Muy-Rangel, M. D., Hernández-Verdugo, S., Villarreal-Romero, M., & Osuna-Rodríguez, J. M. (2016). Fenología reproductiva, rendimiento y calidad del fruto de pitahaya (Hylocereus undatus (How.) Britton and Rose) en el Valle de Culiacán, Sinaloa, México. Agrociencia, 50(1), 61-78.
Silva, A. C., Cavallari, L. L., Sabião, R. R., & Martins, A. B. (2015). Fenologia reprodutiva de pitaya vermelha em Jaboticabal, SP. Ciência Rural, 45(4), 585-590.
Trivellini, A., Lucchesini, M., Ferrante, A., Massa, D., Orlando, M., Incrocci, L., & Mensuali-Sodi, A. (2020). Pitaya, an attractive alternative crop for Mediterranean region. Agronomy, 10, 1065.