
UNED Research Journal (e-ISSN 1659-441X), Vol. 14(2): e4091, December, 2022
ESTUDIO DE CASO
La temperatura y la selección de frutos favorecen la germinación de semillas del árbol tropical Myroxylon balsamum (Fabaceae)
Diego Aguilar-Sandí1![]() & Elmer G. García2
& Elmer G. García2![]()
1. Universidad de Costa Rica, Programa de Posgrado en Biología, San José, Costa Rica; diego.aguilarsandi@ucr.ac.cr
2. Universidad de Costa Rica, Escuela de Biología, San José, Costa Rica; elmer.garcia@ucr.ac.cr
Recibido 26-III-2022 ● Corregido 13-V-2022 ● Aceptado 23-V-2022
DOI: https://doi.org/10.22458/urj.v14i2.4091
|
ABSTRACT. “Temperature and fruit selection favor seed germinations of the tropical tree Myroxylon balsamum (Fabaceae)”. Introduction: Myroxylon balsamum is a tree species native to the Neotropics; and its populations may be declining due to logging and habitat loss. Objective: To evaluate the germinative response of seeds under three temperature conditions, and in fruits selected by seed-chamber thickness. Methods: The fruits were collected in the Central Valley of Costa Rica between November 2019 and June 2020. The germinative response was evaluated at 20ºC, 20/30ºC and 30ºC. The thickness of the seed-chambers was measured and dissected. The fruits were classified as filled (thickness ≥ 0,7cm) and as vain (thickness < 0,7cm). Seed germination within fruits was evaluated at 20ºC, 20/30ºC and 30ºC. Results: As the temperature increased, the time for 50% of the seeds to germinate decreased, and the number of seeds germinated per day, increased. However, there was contamination by pathogens, which caused seed death. Filled fruits germinated well at the three temperatures (92% at 20/30ºC; 86% at 30ºC), while in the vain fruits there was germination only at 20/30ºC (4%) and pathogen contamination could be controlled. Conclusion: The germination for this tree is favored at 20/30ºC and at 30ºC, as long as seeds remain inside fruits with subglobose seed-chambers (thickness ≥ 07cm).
Keywords: balsam tree, Costa Rica, legume, samaras, seed-chamber. |
RESUMEN. Introducción: Myroxylon balsamum es una especie de árbol nativa del neotrópico, cuyas poblaciones podrían estar reduciéndose debido a la tala y a la pérdida de su hábitat. Objetivo: Evaluar la respuesta germinativa de las semillas en tres condiciones de temperatura y en frutos seleccionados según el grosor de la cámara seminífera. Metodología: Los frutos se recolectaron en el Valle Central de Costa Rica entre noviembre de 2019 y junio de 2020. Las semillas se extrajeron y se evaluó la respuesta germinativa a 20ºC, 20/30ºC y 30ºC. Se midió el grosor de las cámaras seminíferas y se diseccionaron. Los frutos se clasificaron en llenos (grosor ≥ 0,7cm) y en vanos (grosor < 0,7cm). Se evaluó la germinación de las semillas dentro de los frutos a 20ºC, 20/30ºC y 30ºC. Resultados: Al incrementarse la temperatura, disminuyó el tiempo para que el 50% de las semillas germinaran y aumentó el número de semillas germinadas por día. Sin embargo, hubo contaminación por patógenos, lo que provocó muerte de semillas. En los frutos llenos, hubo germinación en las tres temperaturas, siendo alta y similar a 20/30ºC (92%) y 30ºC (86%), mientras que en los vanos solo hubo germinación a 20/30ºC (4%). En este caso, la contaminación por patógenos se pudo controlar. Conclusión: La germinación de semillas del árbol de bálsamo se favorece a 20/30ºC y a 30ºC, siempre y cuando permanezcan dentro de frutos que tengan cámaras seminíferas subglobosas (grosor ≥ 0,7cm).
Palabras clave: árbol de bálsamo, Costa Rica, cámara seminífera, leguminosa, sámaras.
|
El árbol de bálsamo, Myroxylon balsamum (L.) Harms (Fabaceae, Papilionoideae), es una especie nativa de los trópicos de América (Sartori et al., 2015). En Costa Rica, se encuentra silvestre en sitios húmedos de la vertiente del Pacífico entre 0 y 850msnm. No obstante, también es posible encontrar árboles cultivados en áreas urbanas del Valle Central, entre 900 y 1200msnm, los cuales son una fuente de germoplasma de fácil acceso (Aguilar-Sandí, 2020a). Los frutos de esta especie son sámaras, en las que se puede distinguir el ala lateral y la cámara seminífera. Esta última estructura contiene, por lo general, una sola semilla reniforme de testa lisa, blancuzca y suave; la presencia de dos semillas en un solo fruto es poco frecuente (Aguilar-Sandí, 2020b).
Desde 1998 se ha considerado que M. balsamum está en peligro de extinción en Costa Rica (Jiménez, 1998). Sin embargo, en 2021 se incluyó en la categoría de Preocupación menor de la Lista Roja de Especies Amenazadas de la Unión Internacional para la Conservación de la Naturaleza (UICN) (Canteiro & Lewis, 2021). Pese a esto, se estima que las poblaciones naturales de los árboles de bálsamo podrían estar experimentando una reducción a nivel global, debido a la tala y a la pérdida de su hábitat natural (Sartori et al., 2015; Canteiro & Lewis, 2021). Esta situación motiva la elaboración de propuestas de investigación que contribuyan con la conservación de esta especie, cuya madera y resina poseen importancia económica (Aguilar-Sandí, 2020a).
Los trabajos publicados sobre germinación de semillas de M. balsamum son escasos. Estos consisten, por una parte, de dos notas técnicas que forman parte de libros de silvicultura, cuya información no es repetible, pues no se muestra la metodología que se empleó para obtenerla [véase Salazar (2000) y Cordero & Boshier (2003)]; y por otra, de dos artículos realizados fuera de Costa Rica, cuyo abordaje metodológico es distinto al desarrollado en la presente investigación [véase Payares-Díaz et al. (2014) y Loayza-Cabezas et al. (2018)]. Adicionalmente, existe una nota técnica publicada en Costa Rica que trata sobre la germinación de semillas de M. peruiferum, la otra especie del género, y no de M. balsamum como pensaron sus autores (véase Rojas-Rodríguez & Torres-Córdoba, 2014).
Los trabajos mencionados anteriormente fueron reunidos y comentados por Aguilar-Sandí (2021). De ese análisis se deduce que, aún no se conoce el efecto de la temperatura en la tasa de germinación de las semillas de esta especie y tampoco se ha establecido una metodología para seleccionar los frutos usados en ensayos de germinación. Tomando esto en cuenta, el objetivo de este estudio es evaluar la respuesta germinativa de las semillas de M. balsamum en tres condiciones de temperatura y en frutos seleccionados según el grosor de la cámara seminífera.
MATERIALES Y MÉTODOS
Recolecta de frutos: Entre noviembre de 2019 y junio de 2020 se recolectaron frutos maduros, directamente del suelo, de dos árboles de M. balsamum. Uno se ubica en el cantón de Santa Ana (09°55'58.6''N, 84°10'42.6''W) y el otro en el cantón de Montes de Oca (09°56'07.3"N, 84°03'05.7"W), provincia de San José, Costa Rica. La zona presenta una estacionalidad climática marcada, con estación seca de diciembre a abril y lluviosa de mayo a noviembre, temperatura promedio mensual de 21 a 24ºC, y precipitación anual de 1580 a 1750mm (Aguilar-Sandí, 2021).
Germinación de semillas en diferentes condiciones de temperatura: Se trabajó con frutos del árbol de Santa Ana que estuvieron almacenados en bolsas plásticas durante cinco meses a aproximadamente 24ºC. Las semillas se extrajeron de los frutos de forma manual; esto se hizo cortando el extremo distal de la cámara seminífera con una tijera de podar. Luego, se seleccionaron aleatoriamente 525 semillas en buen estado (cfr. Aguilar-Sandí, 2020b), se lavaron con agua corriente y se desinfectaron con peróxido de hidrógeno al 3%.
Las semillas se distribuyeron en 21 cajas de Petri de 9,0cm de diámetro con papel filtro (Q3 o Q5) en su interior, y se humedecieron con agua destilada (25 semillas por caja). Posteriormente, las cajas de Petri se distribuyeron en tres recipientes plásticos de 30,0 × 21,5 × 6,5cm con tapa (siete cajas por recipiente) y se colocaron dentro de bolsas plásticas. Esto último se hizo para homogenizar la humedad relativa.
Para los tratamientos de temperatura, se utilizaron tres cámaras de germinación programadas a 20ºC, alternancia 20/30ºC, y 30ºC, respectivamente; todas con un fotoperiodo de 12 horas luz/oscuridad. Dentro de cada cámara se colocó un juego de muestras (i. e. uno de los recipientes plásticos). Transcurridos ocho días, las muestras se rociaron con Agri-mycin 16,5 WP al 0,1% (bactericida) y Amistar al 0,1% (fungicida).
El conteo de semillas germinadas (i. e. cuya radícula había emergido) se realizó cada dos o tres días durante 24 días. Luego, se calculó el porcentaje acumulado final de germinación, el tiempo medio y la tasa de germinación. Las fórmulas utilizadas están disponibles en Aguilar-Sandí (2021). Los promedios (± desviación estándar) obtenidos se compararon con pruebas ANDEVA de una vía y comparaciones múltiples de Tukey realizadas con el programa RStudio (RStudio Team, 2020). Este experimento se realizó en el Centro para Investigaciones en Granos y Semillas (CIGRAS) de la Universidad de Costa Rica (UCR).
Clasificación de los frutos: Se seleccionaron aleatoriamente 100 frutos del árbol de Santa Ana y 100 del árbol de Montes de Oca, y se mezclaron. Seguidamente, se midió el grosor de la cámara seminífera de cada uno con un calibrador. Luego, las cámaras seminíferas se diseccionaron y se determinó el estado de las semillas; para esto, se tomó en cuenta la caracterización de los frutos y de las semillas hecha por Aguilar-Sandí (2020b).
Los frutos se clasificaron en llenos (i. e. con semillas en buen estado) y en vanos (i. e. sin semillas o con semillas dañadas). El promedio (± desviación estándar) del grosor de la cámara seminífera de cada grupo se comparó con una prueba U de Mann-Whitney realizada en RStudio.
Germinación de semillas dentro de los frutos y en diferentes condiciones de temperatura: Se evaluaron frutos procedentes del árbol de Santa Ana que estuvieron almacenados en bolsas plásticas durante cinco meses a 24ºC, aproximadamente. Considerando el resultado de la sección anterior, los frutos se clasificaron en dos grupos según el grosor de la cámara seminífera: llenos (grosor ≥ 0,7cm) y vanos (grosor < 0,7cm).
Se seleccionaron aleatoriamente 150 frutos de cada grupo, se les retiró el ala y se distribuyeron, en partes iguales (N= 50), en tres recipientes plásticos de 30,0 × 21,5 × 6,5cm con tapa y sustrato para germinación húmedo en su interior (turba + agua destilada). Todas las muestras se rociaron con Amistar al 0,1% (fungicida), se taparon y se colocaron dentro de bolsas plásticas.
Posteriormente, las muestras se colocaron en cámaras de germinación con un fotoperiodo de 12 horas luz/oscuridad. Se utilizaron tres cámaras programadas a 20ºC, alternancia 20/30ºC, y 30ºC, respectivamente. Se registró el número de semillas germinadas (i. e. semillas cuya radícula emergió a través del pericarpo) cada dos o tres días durante 35 días. Los porcentajes acumulados finales de germinación se compararon con una prueba Chi cuadrado realizada en RStudio. Este experimento se llevó a cabo en las instalaciones del CIGRAS.
RESULTADOS
Germinación de semillas en diferentes condiciones de temperatura: Las semillas empezaron a germinar al décimo día en los tratamientos de 20/30ºC y 30ºC, mientras que esto ocurrió cinco días después a 20ºC. No se detectó un efecto de la temperatura en el porcentaje acumulado final de germinación, pero sí en el tiempo medio de germinación (Tabla 1) y en la tasa de germinación (Fig. 1). Así, al incrementarse la temperatura, disminuyó el tiempo necesario para que el 50% de las semillas germinaran y aumentó el número de semillas germinadas por día.
A pesar de que se aplicó Agri-mycin 16,5 WP al 0,1% y Amistar al 0,1%, la proliferación de bacterias y hongos no pudo controlarse. Esto provocó muerte de semillas en todos los tratamientos.
Clasificación de los frutos: Los frutos llenos presentaron cámaras seminíferas más gruesas (0,82 ± 0,10cm) que los vanos (0,68 ± 0,15cm) (W= 1798,5; P < 0,001). Esto indica que el grosor de esta estructura puede usarse como criterio cuantitativo para seleccionar frutos que tienen semillas en buen estado y distinguirlos de aquellos que no las tienen o que las tienen dañadas.
TABLA 1
Respuesta germinativa de las semillas de Myroxylon balsamum según la temperatura*
|
Tratamiento |
Porcentaje acumulado final de germinación (%) |
Tiempo medio de germinación (día) |
|
54,00 ± 16,35 A |
18,89 ± 0,75 A |
|
|
20/30ºC |
77,33 ± 22,72 A |
14,69 ± 1,02 B |
|
30ºC |
42,00 ± 40,14 A |
12,85 ± 0,70 C |
*Elaborada con datos de Aguilar-Sandí (2021). Letras distintas indican diferencias significativas según la prueba de Tukey (P < 0,05). Los datos corresponden a promedios ± desviación estándar.
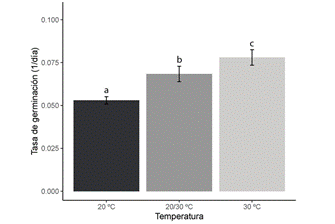
Fig. 1. Tasa de germinación promedio (± desviación estándar) de semillas de Myroxylon balsamum por temperatura. Letras distintas indican diferencias significativas según la prueba de Tukey (P < 0,05). Elaborada con datos de Aguilar-Sandí (2021).
Germinación de semillas dentro de los frutos y en diferentes condiciones de temperatura: En el grupo de frutos llenos (grosor ≥ 0,7cm) hubo germinación en las tres temperaturas; en cambio, en los vanos (grosor < 0,7cm) solo hubo a 20/30ºC, pero el porcentaje de germinación que se obtuvo fue muy bajo en comparación con el obtenido en los llenos en ese mismo tratamiento (χ2= 80,67; g.l.= 1; P < 0,001; Tabla 2). En este último grupo, la respuesta germinativa fue alta y similar a 20/30ºC y 30ºC (χ2= 0,20; g.l.= 1; P= 0,65), mientras que fue baja y distinta a 20ºC (χ2= 51,71; g.l.= 1; P < 0,001; Tabla 2).
Se detectó aparición de hongos en todas las muestras desde el inicio de la prueba, pero la proliferación se controló satisfactoriamente aplicando Amistar al 0,1%. Además, la presencia del pericarpo no impidió que sucediera el proceso de imbibición; tampoco obstaculizó la emergencia de la radícula (Fig. 2A), la elongación del epicótilo (Fig. 2B), ni el desarrollo de la plántula (Fig. 2C).
Germinación y desarrollo de la plántula: La germinación es hipogea y criptocotilar. Los cotiledones son blancuzcos y aumentan de volumen al absorber agua (Fig. 3A, B). En semillas incubadas a 30ºC, la radícula emerge a los 10 o 12 días (Fig. 3C, D). El epicótilo es fotosintético y se elonga rápidamente (Fig. 3E, F). No es posible observar el hipocótilo macroscópicamente.
Las primeras hojas de las plántulas son opuestas e imparipinnadas y tienen cinco folíolos alternos (Fig. 3G). En algunos casos, estas hojas pueden tener filotaxia subopuesta. La yema apical siempre es visible (Fig. 3G); su desarrollo da paso a la formación del tallo y de hojas alternas e imparipinnadas, las cuales tienen un número variable de folíolos (Fig. 4).
Las plántulas conservan los cotiledones hasta por dos meses. Durante ese tiempo, no se observó formación de estípulas ni de yemas laterales.
TABLA 2
Porcentaje acumulado final de germinación de semillas de Myroxylon balsamum dentro de los frutos*
|
Tratamiento |
Frutos llenos (Grosor de la CS** ≥ 0,7cm) |
Frutos vanos (Grosor de la CS** < 0,7cm) |
|
20ºC |
18% A |
0% |
|
20/30ºC |
92% B |
4% |
|
30ºC |
86% B |
0% |
*Elaborada con datos de Aguilar-Sandí (2021). Letras distintas indican diferencias significativas según la prueba Chi cuadrado (P < 0,001). **CS: Cámara seminífera.

Fig. 2. Semilla de Myroxylon balsamum germinando dentro del fruto. A. Emergencia de la radícula; B. Elongación del epicótilo. C. Plántula en desarrollo. (Fotografías de Guillermo Solano).
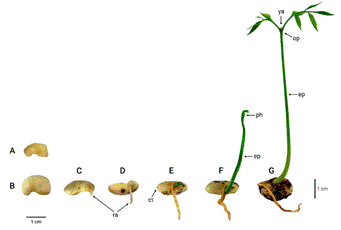
Fig. 3. Germinación y crecimiento de una plántula de Myroxylon balsamum. A. Semilla sin embeber; B. Semilla con 24 horas de imbibición; C. Semilla a los 12 días de iniciada la imbibición (a 30ºC); D. Desarrollo de la radícula; E. Desarrollo del epicótilo; F. Desarrollo del primer par de hojas compuestas; G. Plántula a los 10 días de haber germinado.
Abreviaturas― ct: cotiledones; ep: epicótilo; ph: primeras hojas; op: filotaxia opuesta; ra: radícula; ya: yema apical. [Texto y figura tomados de Aguilar-Sandí (2021, p. 38), CC BY-NC-ND 4.0]

Fig. 4. Plántula de Myroxylon balsamum de dos meses de edad. Abreviaturas― al: primera hoja compuesta alterna;
ct: cotiledones; op: primer par de hojas compuestas opuestas; ya: yema apical.
[Texto y figura de la izquierda tomados de Aguilar-Sandí (2021, p. 39), CC BY-NC-ND 4.0]
DISCUSIÓN
La respuesta de las semillas a las variaciones de temperatura es un rasgo adaptativo asociado con la capacidad de detectar las condiciones favorables para germinar (Copeland & McDonald, 2001; Escobar & Cardoso, 2015). La mortalidad de semillas debida a patógenos impidió visualizar el efecto de la temperatura en el porcentaje de germinación, ya que esto provocó que las desviaciones estándar asociadas a esos promedios fueran muy altas. Sin embargo, el efecto de la temperatura sí pudo observarse en el tiempo medio y en la tasa de germinación. Si bien la germinación se favoreció a 20/30ºC y 30ºC, estas magnitudes no corresponden, necesariamente, a los valores óptimos, dado que para determinar la temperatura óptima se debe evaluar el comportamiento de la tasa de germinación en magnitudes mayores que 30ºC y detectar el punto en el que la tasa decrece (Bewley et al., 2013). No obstante, considerando que hubo un aumento en el número de semillas germinadas por día de 20ºC a 30ºC, cabe inferir que la magnitud óptima es cercana a 30ºC. Respecto a esto, Cordero & Boshier (2003) señalaron que las semillas de M. balsamum germinan entre 25ºC y 35ºC, pero no indicaron el comportamiento de la tasa de germinación en esos valores.
Las diferencias en el grosor de la cámara seminífera de los frutos maduros de M. balsamum se deben, principalmente, al nivel de desarrollo de las semillas en su interior y a la calidad de estas. Aguilar-Sandí (2020b) determinó que los frutos sin semilla tienen la cámara seminífera aplanada, mientras que esta estructura es subglobosa en frutos con semilla. También determinó que, de un total de 800 frutos, 16% no tenían semillas, 59% las tenían en buen estado y 25% las tenían dañadas. Con esto se puso en evidencia la importancia de seleccionar los frutos antes de usarlos en ensayos de germinación. Sin embargo, en dicha investigación no se estableció un criterio cuantitativo para llevar a cabo esa selección; esta carencia se suple aquí.
Salazar (2000) señaló que las semillas de M. balsamum se deben extraer de los frutos; en cambio, Cordero & Boshier (2003) recomendaron no hacerlo, pero en ningún caso se dio una razón que justificara la acción a elegir. Por su parte, Loayza-Cabezas et al. (2018) indicaron que extraer las semillas favorece la germinación, ya que el pericarpo impone latencia física. Sin embargo, esto no coincide con los resultados del presente estudio, pues se comprobó que el pericarpo no es una barrera que limite la germinación ni el subsecuente desarrollo de las plántulas. La baja respuesta germinativa hallada por Loayza-Cabezas et al. (2018) pudo deberse a la presencia de frutos vanos en las muestras evaluadas, puesto que estos autores no implementaron ningún método de selección de frutos.
Tomando en cuenta que las semillas extraídas de los frutos son más propensas al ataque de hongos y bacterias que aquellas que no se extrajeron, cabe suponer que el pericarpo actúa como una cubierta protectora contra patógenos y, en consecuencia, retirarla resultaría contraproducente. No obstante, esta idea debe evaluarse experimentalmente, así como evaluar otros métodos de desinfección, diferentes a los aplicados aquí. En relación con esto, debe señalarse que en ninguno de los trabajos consultados se registró afectación por patógenos ni muerte de semillas (Salazar, 2000; Cordero & Boshier, 2003; Payares-Díaz et al., 2014; Loayza-Cabezas et al., 2018).
En conclusión, la germinación de semillas de los árboles de bálsamo se favorece a 20/30ºC y a 30ºC, siempre y cuando permanezcan dentro de los frutos, los cuales deben tener cámaras seminíferas subglobosas (grosor ≥ 0,7cm), no aplanadas (grosor < 0,7cm).
AGRADECIMIENTOS
Este trabajo es un extracto de la tesis de licenciatura del primer autor (D.A.S.), investigación inédita que se financió con fondos propios y en la que se utilizaron materiales y equipos suministrados por el Centro para Investigaciones en Granos y Semillas (CIGRAS) y por la Escuela de Biología de la Universidad de Costa Rica (UCR).
Durante el desarrollo de la tesis se contó con la valiosa colaboración de Carlos O. Morales, Luis Barboza, Ester Vargas, Guillermo Solano y Andrés Monge. Nuestro más sincero agradecimiento a todos ellos y a los revisores anónimos que examinaron el manuscrito de este artículo.
ÉTICA, CONFLICTO DE INTERESES Y DECLARACIÓN DE FINANCIAMIENTO
Los autores declaramos haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en el manuscrito; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, estamos de acuerdo con la versión final editada del artículo. El respectivo documento legal firmado se encuentra en los archivos de la revista.
La declaración de contribución de cada autor al manuscrito es la siguiente: D.A.S.: Diseño de la investigación, recolección y análisis de datos, redacción del manuscrito y aprobación de la versión final. E.G.G.: Diseño de la investigación, redacción y aprobación de la versión final.
REFERENCIAS
Aguilar-Sandí, D. (2020a). Sinopsis del género Myroxylon (Fabaceae) en Costa Rica. Phytoneuron, 2020,20, 1-10.
Aguilar-Sandí, D. (2020b). ¿Sámaras o frutos samaroides? Estructura macroscópica de los frutos maduros de Myroxylon balsamum (Fabaceae, Papilionoideae). Lilloa, 57(2), 89-98. https://doi.org/10.30550/j.lil/2020.57.2/1
Aguilar-Sandí, D. (2021). Germinación de semillas y crecimiento de plántulas de Myroxylon balsamum (Fabaceae, Papilionoideae). Especie considerada en peligro de extinción en Costa Rica. [Tesis de licenciatura inédita, Universidad de Costa Rica]. Repositorio Institucional de la Universidad de Costa Rica, Kérwá. https://hdl.handle.net/10669/85539
Bewley, J.D., Bradford, K.J., Hilhorst, H.W.M., & Nonogaki, H. (2013). Seeds: Physiology of Development, Germination and Dormancy. Springer Science & Business Media.
Canteiro, C., & Lewis, G. (2021). Myroxylon balsamum. The IUCN Red List of Threatened Species 2021, e.T62026528A62026530. https://dx.doi.org/10.2305/IUCN.UK.2021-2.RLTS.T62026528A62026530.en
Copeland, L.O., & McDonald, M.B. (2001). Principles of seed science and technology (4th ed.). Springer Science + Business Media.
Cordero, J., & Boshier, D.H. (Eds). (2003). Árboles de Centroamérica, un manual para extensionistas. Oxford Forestry Institute/Centro Agronómico Tropical de Investigación y Enseñanza.
Escobar, D.F., & Cardoso, V.J.M. (2015). Germinación y latencia de semillas de Miconia chartacea (Melastomataceae), en respuesta a la luz, temperatura y hormonas vegetales. Revista de Biología Tropical, 63(4), 1169-1184.
Jiménez, Q. (1998). Árboles maderables en peligro de extinción en Costa Rica. Instituto Nacional de Biodiversidad.
Loayza-Cabezas, S., Rodríguez-Trejos, D.A., Hernández-Acosta, E., & Almaráz, J. (2018). Effect of light, temperature and scarification on the germinative capacity and initial growth of Myroxylon balsamum (L.) Harms. Revista Caatinga, 31(2), 415-423.
Payares-Díaz, I., Mario-Contreras, O.A., Medrano-Vélez, M.A., & Millán-Romero, E. (2014). Germinación y desarrollo de plántulas de Myroxylon balsamum (L.) Harms en el departamento de Sucre. Colombia Forestal, 17(2), 193-201. https://doi.org/10.14483/udistrital.jour.colomb.for.2014.2.a05
Rojas-Rodríguez, F., & Torres-Córdoba, G. (2014). Árboles del Valle Central de Costa Rica: reproducción. Chirraca (Myroxylon balsamum). Nota Técnica. Revista Forestal Mesoamericana Kurú, 11(27), 76-78. https://doi.org/10.18845/rfmk.v11i27.1783
RStudio Team. (2020). RStudio: Integrated Development Environment for R. RStudio, PBC. http://www.rstudio.com
Salazar, R. (Coord.). (2000). Manejo de semillas de 100 especies forestales de América Latina. Vol. 1. Centro Agronómico Tropical de Investigación y Enseñanza.
Sartori, A.L.B, Lewis, G.P., Mansano, V.F., & Tozzi, A.M.G.A. (2015). A revision of the genus Myroxylon (Leguminoseae: Papilionoideae). Kew Bulletin, 70, 48. https://doi.org/10.1007/s12225-015-9604-7