
UNED Research Journal (e-ISSN 1659-441X), Vol. 14(supl.1): e3875, June, 2022
Producción de néctar en flores visitadas por colibríes en la Cordillera de Talamanca, Costa Rica
Yandry Gabriela Hernández Barboza1![]()
1. Universidad Estatal a Distancia, Escuela de Ciencias Exactas y Naturales, Proyecto Ecology of Plant and Hummingbird Interactions, 11908 Pérez Zeledón, San José, Costa Rica; yan123hb@gmail.com
Recibido 16-X-2021 Corregido 21-X-2021 Aceptado 25-X-2021
DOI: https://doi.org/10.22458/urj.v14iS1.3875
|
ABSTRACT. “Nectar production in flowers visited by hummingbirds on the Cordillera de Talamanca, Costa Rica”. Introduction: The spatial and temporal variation of the food resource in ecosystems causes discrepancies in the plant-hummingbird functional structure that affect the particular assembly roles. Objective: To evaluate the nectar production and concentration patterns in wild flowers visited by hummingbirds at several elevations over the Cordillera de Talamanca. Methods: Collection of nectar from flowers bagged with a fine mesh for 24 hours at three elevations: high, low and medium. Measurement of flower morphology for each sampled flower, taking temperature, air and soil humidity.Experiments with nectar variations. Results: There is a high variability in nectar production between species and families, associated with the presence of nectar thieves, energy saving strategy and evaporation. At low and medium elevation sites, higher altitudes had lower nectar concentrations, but flowers at higher elevations had more sugary nectar. The smaller flowers produce greater volume and greater concentration. These hummingbirds prefered sweeter nectars, and there was greater visitation at the higher elevation, which is affected by hierarchical behavior and the need for the pollinator. Conclusion: The energy resource for hummingbirds is limited by the morphological barriers between the corollas and the type of beak, the geographic reduction in nectar volume and the competition between species.
Keywords: Specialization, association, secretion, visitation, reabsorption, quantity, quality.
|
RESUMEN. Introducción: La variación espacial y temporal del recurso alimenticio en los ecosistemas causa discordancias en la estructura funcional planta-colibríe que afectan los roles particulares de ensamblaje. Objetivo: Evaluar los patrones de producción y concentración de néctar en flores silvestres visitadas por colibríes en varias elevaciones de la Cordillera de Talamanca. Métodos: Colecta de néctar de flores embolsadas con una malla fina durante 24 horas en tres elevaciones: alta, baja y media. Medición de la morfología floral por cada floral muestreada, toma de temperatura, humedad del aire y del suelo. Experimentos con variaciones en el néctar. Resultados: Se registra una alta variabilidad en la producción de néctar entre especies y familias, asociado a la presencia de ladrones de néctar, estrategia de ahorro de energía y evaporación. En sitios de elevación baja y media, a mayor altitud, se halló menor concentración de néctar, pero las flores de lugares más altos tenían néctar más azucarado. Las flores más pequeñas producen mayor volumen y mayor concentración. Las flores de corola más corta tuvieron mayores volumen y concentración de néctar. Hubo una preferencia de los colibríes hacia néctares más dulces y una mayor visitación en la elevación alta que es afectada por un comportamiento jerárquico y la necesidad del polinizador. Conclusión: El recurso energético para los colibríes es limitado por las barreras morfológicas entre las corolas y el tipo de pico, la reducción en el volumen del néctar y la competencia entre especies.
Palabras clave: Especialización, asociación, secreción, visitación, reabsorción, cantidad, calidad. |
Las interacciones planta-polinizador pueden ser altamente vulnerables al cambio climático debido a las desincronizaciones entre el momento de la floración y la visita de los polinizadores (Maglianesi, 2016), creando un desacople entre las especies que interactúan, un estrés floral y una tensión abiótica. Los colibríes suelen recabar información de cada flor que visitan por medio del forrajeo, especialmente la cantidad y calidad de la recompensa que reciben, para poder estabilizar su presupuesto energético (Bacon et al., 2010). Sin embargo, la capacidad de forrajeo puede ser afectada por la capacidad de respuesta a la variabilidad de los recursos alimentarios en el ambiente, al variar la calidad y la cantidad de néctar, y puede afectar la experiencia previa de los individuos al momento del forrajeo y el estado energético de los individuos (Gómez et al., 2013). El objetivo de la presente investigación fue evaluar los patrones de producción y concentración de néctar en flores silvestres visitadas por colibríes a diferentes elevaciones sobre la Cordillera de Talamanca, Costa Rica, para la generación de información relevante que contribuya con el manejo y conservación de colibríes y las plantas que visitan.
MATERIALES Y MÉTODOS
Los sitios de estudio se localizan en tres bandas altitudinales en la Cordillera de Talamanca: Baja elevación (700msnm): Los Cusingos (9,34035, -83,62899) y Finca Escondida (9,333839, -83,623496) en Quizarrá. Media elevación (1 200 msnm): Centro Biológico Las Quebradas (9,43819,-83,68948). Alta elevación (2 500msnm): Finca Los Encinos en Siberia de Páramo (9,54276, -83,71437).
La metodología empleada se basó en el protocolo de EPHI (2021), para la recolección de néctar, con algunas variaciones como medición floral y la colocación de bebederos artificiales. El periodo de recolección de datos fue entre enero-setiembre del 2020, donde se embolsaron alrededor de 600 flores abiertas visitadas por colibríes con una malla de tela fina a lo largo de los senderos de cada sitio de estudio. Posterior a las 24 horas de estar aisladas, se colectó de cada flor una muestra de néctar con una micropipeta, el líquido extraído fue medido en cm con una regla para la estimación del volumen, usando un refractómetro portátil se determinó la concentración de azúcar en el néctar. En total se embolsaron 111 individuos de 35 especies, colectando néctar de 15 familias a diferentes elevaciones altitudinales. Para cada flor se realizó la medición morfológica, midiendo la longitud y apertura de la corola. Al mismo tiempo se realizó la medición de la temperatura y la humedad del aire usando un termohigrómetro, y un sensor de humedad.
Se utilizó tres bebederos artificiales para analizar las preferencias de especies de colibríes por diferentes concentraciones de néctar en dos elevaciones (alta y baja). Se colocaron 2-3 semanas previo como un período de aprendizaje para los colibríes. Se calculó que los bebederos estuvieran colocados en el bosque por un período de ocho semanas. Se dispusieron los tres bebederos con 10, 20 y 30% de azúcar a un metro entre cada uno, frente a una cámara. El néctar artificial se elaboró con azúcar moreno-cruda. Este néctar se cambió cada 3 días para evitar que se fermente.
RESULTADOS
Características del néctar en flores silvestres visitadas por colibríes: El volumen colectado de néctar varió de 0 a 35μL por flor, mientras que en la concentración de azúcar los números variaron del 0 al 30%. Las especies que presentaron un mayor volumen de néctar por flor fueron Vriesea heliconioides (10,63μL), Centropogon granulosus (10,50μL) y Guzmania donnellsmithii (9,00μL). Mientras que Palicourea padifolia (1,16μL), Bomarea hirsuta (1,25μL) y Hoffmannia dotae (0,94μL) mantuvieron los niveles más bajos. Las especies con el promedio más alto en concentración por flor fueron Passiflora vitifolia con un (30,23%) y Salvia carnea (24,93%). Vriesea heliconioides, Diastema racemiferum y Calathea crotalifera tuvieron las concentraciones más bajas (0-2%).
Entre las familias, Bromeliaceae presentó mayor volumen (8,84μL), seguido por Acanthaceae y Zingiberaceae. Las familias con menor volumen fueron Alstroemeriaceae (1,25μL), Lamiaceae (1,61μL) y Onagraceae (1,87μL). En la concentración la familia que lideró fue Passifloraceae (30,23%), luego Acanthaceae y Lythraceae. Mientras que Marantaceae y Alstroemeriaceae presentaron las concentraciones de néctar más bajas con un 4 y 5%.
Variables abióticas y bióticas que afectan el volumen de néctar: Las variables con efecto en la variación en el volumen fueron la humedad del aire, forma y longitud de la corola. La humedad del aire tiene una relación positiva, aumentando el volumen conforme aumenta la humedad (fig. 1a.). La relación es negativa entre el volumen de néctar y la longitud de la corola, las flores más pequeñas producen mayor volumen (fig. 1b.). En las flores rectas la relación es negativa, donde el volumen por flor es mayor en corolas curvas (fig. 1c).
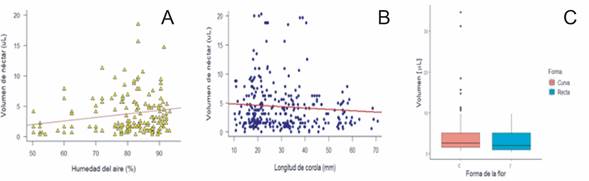
Fig. 1. Relación entre la variable de efectos fijos. (A) Humedad del aire., (B) Longitud de corola. (C)Forma de la flor y el volumen de néctar en los sitios de estudio en la Cordillera de Talamanca, 2020.
Variables abióticas y bióticas que afectan la concentración de azúcar en el néctar de las flores: Las variables abióticas y morfológicas más representativas para explicar la variación en la concentración de sacarosa en el néctar de las flores fueron longitud de la corola, la elevación altitudinal y la humedad del aire, siendo esta última la más significativa. La humedad del aire tiene una relación positiva, conforme aumenta la humedad aumenta la concentración. La elevación baja y media se relaciona de forma negativa con la concentración de néctar, conforme desciende la elevación aumenta la concentración, sin embargo, el comportamiento de la concentración en el néctar cambia en la elevación alta, ya que, la relación entre estas variables es positiva e indica que conforme asciende la elevación asciende la concentración de azúcar (fig. 2b). En la longitud de la corola la relación es negativa, conforma aumenta el tamaño disminuye la concentración (Fig. 2c).
Preferencia de los colibríes a los diferentes niveles de concentración de azúcar en el néctar a dos elevaciones sobre la Cordillera de Talamanca: En los bebederos para colibríes seis especies se acercaron a alimentarse, Panterpe insignis (3 698), Amazilia tzacatl (1 831), Eugenes spectabilis (358), Phaethornis striigularis (306), Lampornis castaneoventris (227) y Phaethornis longirostris (192), siendo Amazilia tzacatl y Panterpe insignis las especies alfa. Para cada concentración de azúcar, el néctar artificial con 30% de sacarosa fue el más seleccionado por los colibríes con 4 769 interacciones, seguido por el de 20% (1 385) y de último el de 10% con solo 449 visitas. Además, se observó una relación significativa entre los bebederos de la elevación alta y el néctar artificial más concentrado (Fig. 3).
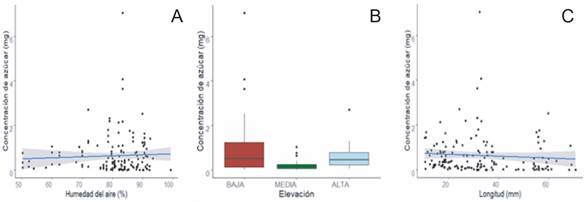
Fig. 2. Relación positiva entre la variable de efecto fijo. (A) Humedad del aire, (B) Elevación, (C) Longitud y la concentración de azúcar en los sitios de estudio de la Cordillera de Talamanca, 2020. Nota: las líneas horizontales representan la mediana, las cajas indican los percentiles 25% y 75%, los bigotes indican el rango de los datos y los puntos son valores atípicos encontrados en la base de datos.
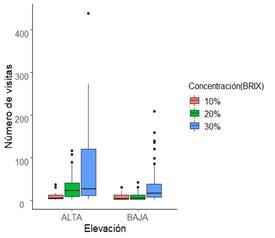
Fig. 3. Preferencia de los colibríes hacia diferentes niveles de concentración de azúcar en cada sitio de estudio en las dos elevaciones altitudinales.
DISCUSIÓN
Características del néctar en flores silvestres visitadas por colibríes: El volumen y la concentración de azúcar en el néctar de las flores varió entre especies y familias. Esta variación puede ser resultado de una serie de especializaciones desarrolladas por la planta para ahorrar energía. Ejemplo, flores con grandes cargas de polen, suelen reducir el costo asociado a producir néctar para contener este recurso energético en la producción de semillas. La cantidad de néctar puede variar cuando las plantas regulan sus picos de floración de acuerdo con el aumento de individuos de colibríes. La temporada de floración y el número de visitas de los polinizadores puede causar la variedad en la producción de néctar en especies y familias, haciendo que cada planta adapte la producción de néctar de acuerdo con las necesidades de sus polinizadores (Deliso, 2007). Así mismo, las plantas pueden absorber el néctar cuando no existe visitación por colibríes y contener este recurso para visitas legitimas, evitando el robo de néctar (Corbet, 2003).
Variables abióticas y bióticas que afectan el volumen de néctar: La fuerte relación entre cantidad de néctar producido y la humedad del aire puede ser explicado por el estrés hídrico en el que crece la planta, al estar obligada a secretar agua para equilibrar los efectos de la evaporación, y así encontrar mayor volumen de néctar en las flores, aunque no precisamente más concentrado. Entre más alto sea el vapor de agua en la atmósfera, más volumen de néctar producido, por el almacenamiento de agua en los nectarios (Corbet, 2003).
La variación en la morfología de la corola entre flores de diferentes especies de plantas puede ser explicada por la interacción entre planta-polinizador, en donde las flores adaptan sus corolas de acuerdo con su visitante polinizador en respuesta a una especialización. La forma de la corola influye en el uso del recurso energético como una barrera, evitando una competencia interespecífica entre colibríes de diferentes tipos de picos (Maglianesi et al., 2014), esto favorece la acumulación de néctar y reduce su exposición a la evaporación (Corbet, 2003; McDade & Weeks, 2004), beneficiando las flores con corolas largas y curvas que protegen las cámaras de néctar. En aquellas flores pequeñas y rectas se pierde mayor néctar, al ser expuestas por robadores de néctar, que disminuyen la cantidad y calidad del néctar y provoca una menor tasa de visitación por colibríes, afectando la producción de semillas (Salinas-Esquivel et al., 2013).
Variables abióticas y bióticas que afectan la concentración de azúcar en el néctar de las flores: La humedad del aire tiende a tener un comportamiento sobre la concentración similar al comportamiento observado en el volumen de néctar, en donde conforme asciende la humedad, aumentan los niveles en la concentración de azúcar. Corbet (2003) explica que la concentración de néctar puede variar según la hora del día, así estas especies presentan un néctar menos concentrado por la noche cuando la humedad relativa es baja, y aumenta por la mañana cuando la humedad es alta. La reducción del agua cierra los estomas y detiene la fotosíntesis, necesaria para la producción de néctar provocando una variación en la concentración de azúcar de un día a otro (Deliso, 2007).
La elevación altitudinal tiene un efecto en la concentración de azúcar. Así, el néctar resultó más concentrado en la elevación baja, disminuyendo considerablemente en la elevación media, y aumentando la concentración en la elevación alta. Explica Descamps et al. (2018) que conforme asciende la temperatura (elevaciones bajas), aumenta la producción de néctar, hasta llegar a un máximo de temperatura en la que el néctar empieza a reducir su volumen por los efectos del estrés hídrico. El volumen de néctar asciende de nuevo cuando se reduce este estrés y la planta puede producir néctar nuevamente, caso observado en la elevación alta. Las plantas en la elevación alta deben de producir néctar más concentrados para satisfacer las necesidades de los colibríes que en esos ecosistemas se encuentran (Clasen et al., 2019).
Se observa una relación negativa entre la longitud de la corola y la concentración del néctar, es decir, flores más largas poseen un néctar menos concentrado. Entre más larga sea la corola se evita la pérdida de agua y el néctar se vuelve menos concentrado. Lo opuesto ocurre con las corolas pequeñas donde el agua del néctar tiene más posibilidad de evaporarse y concentrar los gramos de sacarosa.
Preferencia de los colibríes a los diferentes niveles de concentración de azúcar en el néctar a dos elevaciones sobre la Cordillera de Talamanca: No todas las especies de colibríes logran identificar los bebederos como un recurso alimenticio por sus características morfológicas, al no ser tractivas (Maglianesi et al., 2014). Los colibríes que realizaron visitas tuvieron una preferencia marcada hacia los bebederos con mayor concentración de azúcar (20% y 30% respectivamente). Estas visitas pueden depender de la conducta territorial de las especies alfa, que defienden territorios con mayor recurso energético (Márquez-Luna et al., 2017). Una mayor preferencia por néctares más concentrados en los bebederos podría explicarse por el hecho de que las especies de colibríes en elevaciones altas necesitan un recurso energético en mayor cantidad y calidad, esto para contrarrestar las situaciones climáticas severas que enfrentan como bajas temperaturas y poca floración. Según Avalos (2019) la presencia de los colibríes en los bebederos puede responder según la situación de disponibilidad de recursos energéticos, siendo mayor en la época lluviosa que en la época seca debido a la abundancia de flores en este periodo.
AGRADECIMIENTOS
Agradezco a todos los colaboradores del proyecto Ecology of Plants and Humminbirds Interactions (EPHI) por la asesoría, financiamiento y planificación para la presente investigación, en especial a María Maglianesi por ser directora de tesis.
ÉTICA, CONFLICTO DE INTERESES Y DECLARACIÓN DE FINANCIAMIENTO
Declaro haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la preparación de este documento; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, estoy de acuerdo con la versión editada final de esta publicación. El respectivo documento legal firmado se encuentra en los archivos de la revista.
REFERENCIAS
Avalos, G. (2019). Recomendaciones para mejorar el manejo de comederos artificiales para colibríes. Zeledonia, 23(2), 69-75. https://bit.ly/3Do4iIU
Bacon, I., Hurly, A., & Healy, S. (2010). Both the past and the present affect risk-sensitive decisions of foraging rufous hummingbirds. Behavioral Ecology, 21(1), 626–632. https://doi:10.1093/beheco/arq031
Clasen, A., Eardley, C., Hemp, A., Peters, M., Peters, R., Ssymank, A., & Steffan, I. D. (2019). Specialization of plant–pollinator interactions increases with temperature at Mt. Kilimanjaro. Ecology and Evolution, 10, 2182-2195 https://doi.org/10.1002/ece3.6056
Corbet, S. A. (2003). Nectar sugar content: estimating standing crop and secreation rate in the field. Apidologie, 34(1), 1-10. https://doi:10.1051/apido:2002049
Deliso, E. (2007). Climate Change and the Hummingbirds of the Monteverde Cloud Forest, Costa Rica (1 ed.). Centro Científico Tropical. https://bit.ly/2YSNOd0
Descamps, C., Quinet, M., Baijot, A., y Jacquemart, A. (2018). Temperature and water stress affect plant–pollinator interactions in Borago officinalis (Boraginaceae). Ecology and Evolution, 8(6), 3443-3456. https://doi.org/10.1002/ece3.3914
EPHI Ecology of Plant and Hummingbird Interactions (2021). Research and Study Approach for the project Developing the predictive ecology of plant-animal interactions across space and time. https://hummingbird.bio/study-approach/
Gómez, L. R., Ortiz-Pulido, R., & Lara, C. (2013). Sensibilidad al riesgo durante el forrajeo en los colibríes Hylocharis leucotis y Selasphorus platycercus. Revista Mexicana de Ornitología, 14(1), 7-16. https://bit.ly/3Dk7sNP
Maglianesi, M. (2016). Efectos del cambio climático sobre la polinización y la producción agrícola en América tropical. Revista Ingeniería, 26(1), 11-20. https://bit.ly/3qEXao5
Maglianesi, M., Blüthgen, N., Böhning-Gaese, K., y Schleuning. M. (2014). Morphological traits determine specialization and resource use in plant–hummingbird networks in the neotropics. Ecology, 95(12), 3325-3334. https://bit.ly/30q65PL
Márquez-Luna, U., Lara, C., & Ortiz-Pulido, R. (2017). La calidad del néctar afecta la conducta territorial y de forrajeo en hembras del colibrí lucifer (Calothorax lucifer): Un experimento. Ornitología Neotropical, 26(1), 67-75. https://core.ac.uk/reader/288303490
McDade, L. A., & Weeks, J. A. (2004). Nectar in Hummingbird-pollinated Neotropical Plants I: Patterns of Production and Variability in 12 Species. Biotropica, 36(2), 192-215. https://www.jstor.org/stable/30043108
Salinas-Esquivel, L., Arizmendi, M. del C., Domínguez, C., Castillo-Guevara, C., & Lara, C. (2013). Producción De Néctar y Flores como Rasgos Compensatorios al Robo de Néctar en Bouvardia Ternifolia (Rubiaceae): Experimentos de Campo. Botanical Sciences, 91(1), 85-92. http://www.scielo.org.mx/pdf/bs/v91n1/v91n1a7.pdf