
Morfometría corporal y reproducción de la rata de campo Sigmodon hirsutus (Rodentia: Cricetidae) en Costa Rica
Javier Monge1 ![]()
1. Universidad de Costa Rica, Escuela de Agronomía, Centro de Investigación en Protección de Cultivos, Instituto de Investigaciones Agrícolas, Facultad de Ciencias Agroalimentarias, San José, Costa Rica; javier.monge@ucr.ac.cr,
Recibido 8-X-2019 Corregido 17-II-2020 Aceptado 20-II-2020
DOI: https://doi.org/10.22458/ urj.v12i1.2887
|
ABSTRACT. “Body morphometry and reproduction of the field rat Sigmodon hirsutus (Rodentia: Cricetidae) in Costa Rica”. Introduction: The rat (Sigmodon hirsutus) is common in agricultural environments in Central America. Of this species little biological and ecological information is available, given its recent reclassification as a species. Objective: To describe the body morphometry, used in turn as a criterion of body condition and to determine your reproductive cycle of the field rat. Methods: The study was carried out in the canton of La Cruz, Province of Guanacaste, Costa Rica from March 2009 to April 2010. I did 29 fortnightly samplings of three consecutive nights on organic pineapple crops. I measured the length and weight of individuals to establish a relationship between these variables and use them to estimate the body condition index. I determined the reproductive cycle based on the state of pregnancies, presence of juveniles, and in complementary form the testicular position in males. By body characteristics and reproductive activity, age categories were defined. Results: I collected 418 specimens of S. hirsutus, for a sex ratio of 1:0.6 (male:female). Males are bigger and heavier. The body condition index (mm/g) ranged from 1 to 6,4 for males and between 1 and 8,3 for females, with an inverse relationship to the weight category. Of the 154 females collected, 36,4% were pregnant, all of which were present in all months and most often in April 2009, June and August. The presence of juveniles was greatest in April 2009 and June, although they were present in almost every month of the year. The scrotal position of testicles was the most common (54,4%). The age categories being represented throughout the year, suggesting a reproductive activity for the species and site throughout the year. Conclusion: The rat S. hirsutus is of medium size (141,65mm and 84,13g and 131,81mm and 69,07g, for males and females, respectively), and presents sexual dimorphism. Reproductive activity occurs throughout the year.
Keywords: d Rodents, reproduction, morphology, pineapple, Costa Rica.
|
RESUMEN. Introducción: La rata de campo (Sigmodon hirsutus) es común en ambientes agrícolas de América Central, de la cual se dispone de relativa poco información biológica y ecológica, dada su reciente reclasificación como especie. Objetivos: Describir la morfometría corporal, utilizada a su vez como criterio de condición corporal y determinar su ciclo reproductivo de la rata de campo. Métodos: El estudio se realizó en el cantón La Cruz, provincia de Guanacaste, Costa Rica entre marzo de 2009 a abril de 2010. Se realizaron 29 muestreos quincenales de tres noches consecutivas en cultivos de piña orgánica. Se tomaron medidas de longitud y peso de los individuos para establecer una relación entre estas variables y utilizarlas para estimar el índice de condición corporal. Se determinó el ciclo reproductivo basado en el estado de preñez, presencia de juveniles, y en forma complementaria la posición testicular en los machos. Por características corporales y actividad reproductiva se definió categorías de edad. Resultados: Se colectaron 418 especímenes de S. hirsutus, para una proporción de sexos de 1:0,6 (machos:hembras). Se presenta un dimorfismo sexual con un mayor peso y tamaño para los machos. El índice de condición corporal (mm/g) osciló entre 1 y 6,4 para los machos y entre 1 y 8,3 para las hembras, con una relación inversa con la categoría de peso. De las 154 hembras colectadas, el 36,4% se encontraban preñadas, las cuales presentes en todos los meses y con mayor frecuencia en abril de 2009, junio y agosto. La presencia de juveniles fue mayor en los meses de abril de 2009 y junio, aunque estuvieron presentes en casi todos los meses del año. La posición escrotal de testículos fue la más común (54,4%). Las categorías de edad presentes a través del año, sugieren una actividad reproductiva para la especie y sitio durante todo el año. Conclusión: La rata S. hirsutus es de tamaño mediano (141,65mm y 84,13g y de 131,81mm y 69,07g, para machos y hembras, respectivamente), y presenta dimorfismo sexual. La actividad reproductiva se presenta en todo el año.
Palaras clave: Roedores, reproducción, morfología, piña, Costa Rica.
|
La rata de campo (Sigmodon hirsutus) es una especie común en pastizales y áreas de producción agrícola de Centroamérica y la parte norte de Suramérica. Esta especie fue considerada como una subespecie de S. hispidus, hasta que estudios de Peppers, Carroll y Bradley (2002), Carroll y Bradley (2005) permitieron reclasificarla como especie. Por lo tanto, la información proveniente de estudios realizados en sitios en Centroamérica y parte norte de Suramérica corresponde a S. hirsutus, aunque la refieran como S. hispidus. Si bien existe información sobre esta especie, la misma es limitada y de algunos aspectos de su biología se dispone de datos aislados o basados en estudios de corta duración.
Con respecto a la reproducción de esta especie, se considera que aparentemente se reproduce durante todo el año (Fleming, 1970; Vivas & Calero, 1985). A su vez, la presencia de hembras preñadas y jóvenes indica que se reproducen al menos durante los meses de octubre a enero, en un estudio que abarcó de agosto a enero (Monge, 2008). En cuanto al tamaño de camadas, se ha estimado entre 2,8 y 5,7 (Bowdre, 1971), así como en 6,25 ± 1,5 embriones por camada (Monge, 2008).
Relacionada con la reproducción está la edad de los individuos, aspecto del que se carece de criterios para la clasificación de individuos de esta especie, aun cuando Fleming (1970) se refiere a categorías de edad (jóvenes, subadultos y adultos) basado en la condición del pelaje, sin detallar el criterio. A su vez, Monge (2008) propuso una clasificación arbitraria al considerar como juveniles a aquellos individuos cuyo peso correspondiera al 20% menor del ámbito de pesos (20-206g) de una muestra 39 individuos, subadultos de 21-50% y adultos más del 50%, por lo que individuos con peso igual o inferior a 57,2 g se clasificaban como juveniles, de 57,3 a 113g como subadultos y mayor de 113 como adultos. Estos valores guardan cierta similitud con la clasificación propuesta por Odum (1955) para S. hispidus al considerar que los individuos con peso inferior a 60g como juveniles, de 60-110g como adultos jóvenes y aquellos con peso superior a 110g como adultos viejos. A su vez, Layne (1974) utilizó para esa misma especie los límites de peso de menor a 50g (jóvenes), 50-100g (subadultos) y más de 100g (adultos).
Otro aspecto relacionado con la reproducción es la estructura poblacional, es decir la proporción de sexos, así como la de las categorías de sexo-edad de los individuos que componen la población. Existe una tendencia de más capturas de machos que hembras, aunque no se evidencia una diferencia significativa a la relación 1:1 (Vivas & Calero, 1985; Monge, 2008).
La relación entre el peso y longitud corporal, permite por medio de modelos predecir una u otra variable, y a su vez, estimar la condición de cada individuo en función del peso mostrado comparado con el esperable según el modelo (Krebs & Singleton, 1993). Vivas y Calero (1985) utilizaron un índice de condición corporal (longitud/peso) para S. hispidus (actualmente S. hirsutus) y encontraron oscilaciones durante el año, sin que se presentara una relación con variables climáticas, salvo en los juveniles que muestra una cierta relación inversa con la temperatura, aunque no significativa.
El propósito de este documento es describir la morfometría corporal, utilizada a su vez como criterio de condición corporal y determinar el ciclo reproductivo de la rata de campo (Sigmodon hirsutus) en un área de cultivo de piña de producción orgánica.
MATERIALES Y MÉTODOS
El estudio se realizó en Santa Cecilia, cantón La Cruz, provincia de Guanacaste, Costa Rica (11°03ʹ36ʺN & 85°24ʹ58ʺO) a una elevación de 334msnm, durante el período de marzo de 2009 a abril de 2010. El sitio durante los 14 meses de estudio tuvo una precipitación de 2 583mm, la máxima temperatura registrada fue de 37°C y la mínima de 17°C, con dos estaciones climáticas, la lluviosa que se extendió entre los meses de enero a abril, con precipitaciones mensuales entre los 18 y 62mm, y la lluviosa de mayo a diciembre con precipitaciones mensuales entre 124 y 474mm.
Se realizaron 29 muestreos quincenales de tres noches consecutivas que iniciaban a las 15:00 horas y terminaba a las 7:00 horas del día siguiente. Las capturas se realizaron con trampas Victor (Woodstream Corp., Lititz, PA, USA) en ubicación fija alrededor de siete lotes de piña con un distanciamiento de 10m entre trampas, para un total de 134 trampas por muestreo. Se utilizó como cebo atrayente un trozo de piña. La producción de piña en el sitio fue de tipo orgánica, por lo que no se utilizaban productos químicos para el control de los roedores u otros organismos.
Los individuos capturados fueron congelados para su posterior análisis. Para la toma de datos, los individuos fueron descongelados y se tomaron las medidas de largo total y de la cola, para luego estimar la longitud corporal (largo total menos el largo de cola), se pesaron y se determinó su sexo. En los machos se observó la posición de los testículos (abdominal o escrotal), y a las hembras se les hizo una incisión abdominal para extraer el tracto reproductivo para conocer su tamaño, así como para determinar su estado reproductivo. En el caso de las hembras preñadas se contabilizó los embriones.
Los individuos fueron clasificados por categorías de peso, según las siguientes clases: menor o igual a 30g, 31-60g, 61-90g, 91-120g, 121-150g, 151-180g y más de 180g. Se estableció una relación entre peso y longitud corporal y se determinó la condición corporal basada estas variables, según la categoría de peso y sexo.
El ciclo reproductivo se determinó según el estado de preñez, así como la presencia de individuos jóvenes través del período de estudio. La posición testicular de los machos se usó como referencia complementaria. Para determinar la estructura poblacional, es decir la proporción de sexo y edad, se usó las categorías de peso como referencia de la edad.
Ética, conflicto de intereses y declaración de financiamiento: El autor declara haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en el manuscrito; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, está de acuerdo con la versión editada final del documento. El respectivo documento legal firmado se encuentra en los archivos de la revista.
RESULTADOS
Durante el periodo de estudio se colectaron 418 especímenes de S. hirsutus, de los cuales 264 eran machos y 154 hembras, para una proporción de sexos de 1:0,6 (machos:hembras), la que difiere de una relación de igualdad (chi cuadrado= 28,95, p< 0,001). Las capturas por muestreo oscilaron entre 5 y 29 individuos, con representación de ambos sexos en todos los muestreos (Fig. 1). La proporción de sexos por muestreo osciló entre 1:0,1 a 1:1,5.
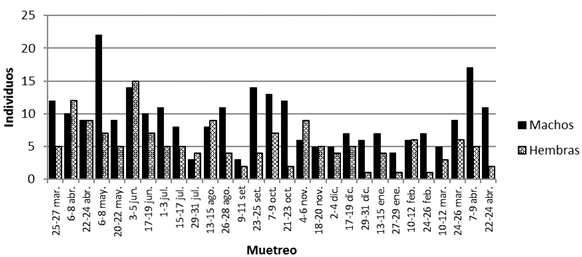
Fig. 1. Capturas de machos y hembras de S. hirsutus por muestreo.
El peso promedio ± desviación estándar de los machos fue de 84,13 ± 44,30g (13-206g), mientras que para las hembras fue de 69,07 ± 38,85g (9-172g), evidenciándose una diferencia significativa (z= 3,61; p< 0,001). En cuanto a lo longitud corporal, lo machos tuvieron un promedio de 141,65 ± 28,36mm (71-216mm) y las hembras de 131,81 ± 26,54mm (75-187mm), existiendo una diferencia significativa (z= 5,26; p< 0,001). De esta manera, se presenta un dimorfismo sexual con un mayor peso y tamaño para los machos.
Al clasificar a los individuos por categorías de peso, se obtuvo que las categorías más representadas, fueron las de 61-90g y 91-120g, con 23,1 y 22,8%, respectivamente, seguida de la de 31-60g con un 21,6% y luego la de peso ≤30g, con un 15,8% (Cuadro 1). La proporción de sexos por categoría de peso osciló entre 1:0,3 a 1:1,1 (machos:hembras), presentándose la situación más extrema en la última categoría de peso (Mas de 180g) representada solo por machos. En las categorías de peso de ≤30g y de 31-60g no se evidencia una diferencia significativa a la relación de igualdad de proporción de sexos (chi-cuadrado = 0,75 y 0,10, respectivamente, p<0,05), mientras que en las siguientes categorías hay una diferencia significativa con mayor representación de machos.
CUADRO 1
Composición de la población de S. hirsutus por categoría de peso
|
Categoría de peso (g) |
Machos |
Hembras |
Total |
Prop. Machos |
Prop. Hembras |
Prop. Total |
|
≤ 30 |
36 |
29 |
65 |
13,85 |
19,21 |
15,78 |
|
31-60 |
43 |
46 |
89 |
16,54 |
30,46 |
21,60 |
|
61-90 |
71 |
24 |
95 |
27,31 |
15,89 |
23,06 |
|
91-120 |
57 |
36 |
94 |
22,92 |
23,84 |
22,82 |
|
121-150 |
32 |
12 |
44 |
12,31 |
7,95 |
10,68 |
|
151-180 |
15 |
4 |
19 |
5,77 |
2,65 |
4,61 |
|
>180 |
6 |
0 |
6 |
2,31 |
0,00 |
1,46 |
|
Total |
260* |
151* |
412 |
100 |
100 |
100 |
|
* La diferencia de datos de machos y de hembras con respecto a la totalidad de la muestra de estudio, se debe a individuos con información incompleta, ya sea de peso o de longitud. |
||||||
En la Fig. 2 se muestra la estructura poblacional de S. hirsutus en donde se aprecia la representación para las categorías de peso y sexo. Se observa la tendencia de una mayor representación de machos para la mayoría de las categorías de peso, excepto para la categoría de 31-60g, aunque no existe una diferencia significativa a la relación de igualdad.
Se estableció la relación entre el peso y la longitud corporal de los especímenes capturados, la cual es definida por la ecuación L = 41,63ln(P) - 35,92, según un modelo logarítmico (R²= 0,942) (Fig. 3). A su vez, para estimar el peso basado en el valor de longitud se puede utilizar la ecuación P= 2,870e0,022L según un modelo exponencial (R²= 0,942), donde L es la longitud (mm) y P el peso (g).
El índice de condición corporal basada en la relación entre la longitud y el peso corporal (mm/g) para cada individuo, osciló entre 1 y 6,4 para los machos y entre 1 y 8,3 para las hembras (Fig. 4). Se puede observar que conforme aumenta la categoría de peso, se disminuye el valor de índice de condición corporal.
En cuanto a la condición reproductiva, el criterio más confiable es el estado de preñez de las hembras, dado que este estado representa un aporte real a la población, más allá que alguna sugerencia de actividad o inactividad reproductiva, pero sin certeza de participación en la reproducción. De las 154 hembras colectadas, 55 se encontraban preñadas, lo que representa el 36,4%. Las hembras con muestras de inicio de preñez, dado el abultamiento de sus cuernos uterinos, así como en evidente estado de preñez con desarrollo de embriones, pesaron entre 51 y 172g, es decir que se ubican entre la segunda categoría de peso (31-60 g) y la sexta (151-180g). En la primera categoría de peso (menos de 30g) no se obtuvo hembras con muestras de preñez, además tenían el tracto reproductivo muy pequeño y por las características de un pelaje sedoso puede asumirse que esta clase está compuesta por individuos juveniles. La segunda categoría de peso (31-60g) incluyó cuatro hembras con muestras de inicio de preñez con pesos de 51, 57, 58 y 60g, cuyo tracto reproductivo de las hembras de esta categoría es relativamente pequeño, por lo que podría considerarse una categoría de edad de subadultas como una transición entre juveniles y adultas. La proporción de hembras preñadas del total de hembras se incrementó en cada categoría de peso, así en la clase cuyo peso es igual o menor a 30g (juveniles) no hubo hembras preñadas, en la clase 31-60g (subadultas) el 8,3% presentan muestras de inicio de preñez, y en las siguientes categorías de peso se presentan hembras preñadas en proporciones de 37,5 % (61-90g), 75,7 % (91-120g), 83,3% (121-150g) y del 100% (151-180g) (Cuadro 2).
Las hembras preñadas estuvieron presentes en todos los meses del año, con una mayor presencia en los meses de abril de 2009 y junio con 8 hembras cada uno y agosto con seis hembras. En la Fig. 5 se muestra la cantidad de capturas de hembras de S. hirsutus preñadas y no preñadas, sin incluir individuos de peso igual o inferior a 30g, considerados como juveniles. Si bien no se aprecia una tendencia de acumulación de mayor cantidad de hembras preñadas en alguna época de año, en algunos meses hay una importante proporción de hembras preñadas que igualan o superan el 50% del total de hembras adultas y subadultas, como en los meses de abril de 2009, agosto, setiembre, diciembre, febrero, marzo de 2010 y abril de 2010, entre cuyos meses se encuentran otros con una menor proporción de hembras preñadas.
En cuanto a tamaño promedio de camada fue de 4,4 ± 1,3 (1-7) embriones y osciló entre 3,5 a 4,8 según categoría de peso (Cuadro 2). El tamaño promedio de camada por mes osciló entre 3 y 6, con un tamaño superior al inicio del estudio, con un leve descenso entre los meses de mayo y junio, para mantenerse luego con oscilaciones con tamaño promedio entre 4 y 5 embriones por camada (Fig. 6). El tamaño de camada más frecuente fue de 4 (25 hembras), seguido de 5, 6 y 3, con 9, 7 y 3 hembras, respectivamente.
|
|
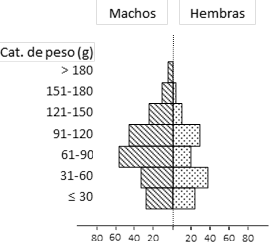
Fig. 2. Estructura poblacional de S. hirsutus
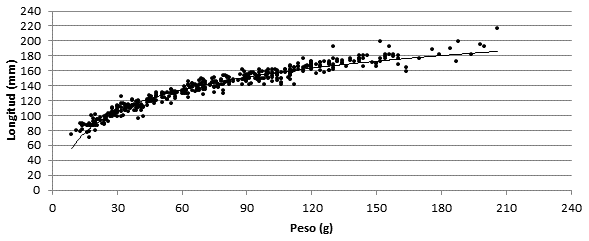
Fig. 3. Relación entre peso y longitud corporal de S. hirsutus
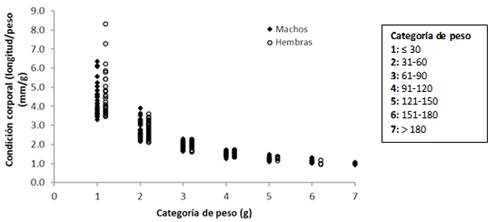
Fig. 4. Condición corporal de S. hirsutus por categoría de peso
CUADRO 2
Cantidad de hembras de S. hirsutus, condición reproductiva y tamaño de camada
|
Categoría de peso |
Hembras |
Preñadas |
Tamaño de camada |
|
≤ 30 |
29 |
0 |
- |
|
31-60 |
46 |
4 |
3,5 ± 1,0 |
|
61-90 |
24 |
9 |
3,3 ± 1,5 |
|
91-120 |
36 |
28 |
4,5 ± 1,1 |
|
121-150 |
12 |
10 |
4,7 ± 1,3 |
|
151-180 |
4 |
4 |
4,8 ± 1,3 |
|
Total |
151 |
55 |
4,4 ± 1,3 |
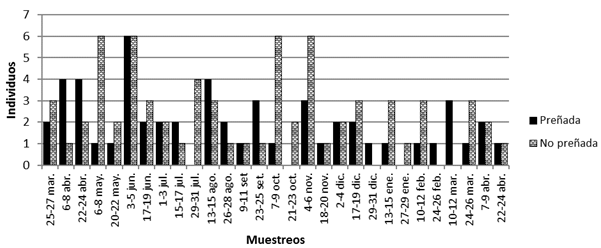
Fig. 5. Capturas de hembras de S. hirsutus preñadas y no preñadas
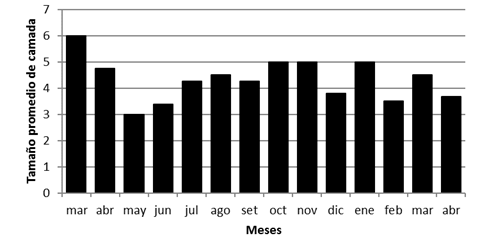
Fig. 6. Tamaño promedio de camada de S. hirsutus
En cuanto a los machos, la condición de testículos sugiere una tendencia de posición abdominal (T.A.) en las categorías de menor peso, dado que la totalidad de los individuos de la primera categoría de peso (≤30g), así como el 79% de los de la categoría de 31-60g y el 18% de los individuos de la siguiente categoría (61-90%) tenían testículos en posición abdominal. Los machos de la categoría de peso menor carecían de escroto, obviamente sin posibilidad de tener testículos con posición escrotal, lo que complementa lo indicado para las hembras que esta categoría está representada por individuos juveniles y sin capacidad de reproducirse.
Los individuos con posición testicular intermedia, llamada semiescrotal (T.S.E.) representan solo al 8,7% del total de machos, ubicados en las categorías de peso 31-60g (21,7%), 61-90g (73,9%) y 91-120g ( 4,3%). La posición escrotal de testículos fue la más común, observada en el 54,4% de los machos capturados, presentes en casi todas las categorías de peso, excepto en la primera (≤ 30g) y con una limitada representación en la segunda categoría de peso (31-60%), ya que de esta categoría solo el 7% tienen esta posición testicular. Para las siguientes categorías de peso, representa la posición de testículos predominante, con un 56,9% (61-90g), 89,7% (91-120g), 87,5% (121-150g), 93,3% (151-180g) y 100% para la categoría de más de 180 g de peso corporal (Fig. 7). No se observó una tendencia de predominio de individuos para cada posición testicular a través del tiempo, sino que tienden a estar presente en todos los muestreos, salvo para la posición semiescrotal que al ser menos común, en algunos muestreos no estuvo representada.
La presencia de jóvenes, considerando ambos sexos, tendió a acumularse en los meses de abril de 2009 y junio aunque estuvieron presentes en las colectas hasta el mes de setiembre, presentándose un período entre setiembre y enero con ausencia de individuos con peso igual o inferior a los 30g, salvo en noviembre y diciembre en que se colectaron tres individuos (Fig. 8).
La presencia de diferentes categorías de peso a través del año, las que pueden relacionarse por categorías de edad (juveniles, subadultos y adultos) (Fig. 9) sugieren de una actividad reproductiva durante todo el año, con épocas de mayor actividad definidas por mayor presencia de juveniles y proporción de hembras preñadas con respecto a cantidad de hembras subadultas y adultas. Así, la mayor presencia de hembras preñadas entre abril y agosto y de juveniles en abril de 2009 y junio, indican que de abril a agosto fue un periodo propicio para la reproducción de la especie en el sitio de estudio.
Si se consideran estas tres categorías de edad, los ámbitos de peso y longitud para machos y hembras corresponde a como se muestra en el Cuadro 3.
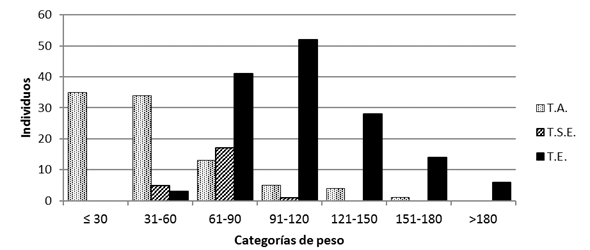
Fig. 7. Posición testicular de S. hirsutus según categoría de peso
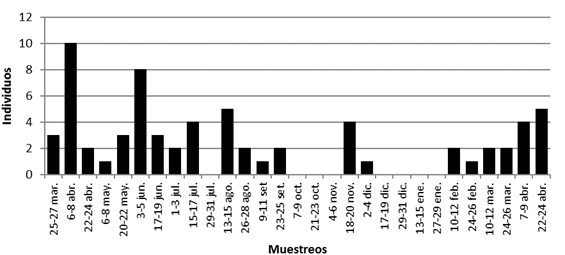
Fig. 8. Capturas de jóvenes de S. hirsutus
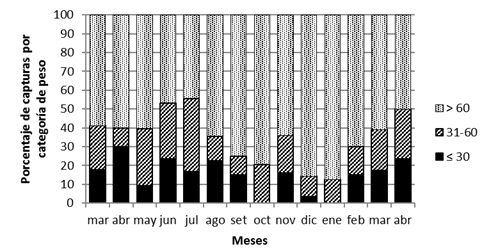
Fig.9. Proporción de individuos de S. hirsutus por categorías de peso
CUADRO 3
Peso y longitud por categoría de peso (edad) de S. hirsutus
|
Peso promedio ± D.E. y ámbito de peso (g) |
||||||
|
Peso (g) |
n |
Machos |
n |
Hembras |
n |
Ambos |
|
≤ 30 (Jóvenes) |
36 |
22,14 ± 5,39 13 - 30 |
29 |
21,90 ± 6,34 9 - 30 |
65 |
21,90 ± 6,34 9 - 30 |
|
31-60 (Subadultos) |
43 |
44,05 ± 8,71 32 - 60 |
46 |
44,15 ± 8,36 31 - 60 |
89 |
44,15 ± 8,36 31 - 60 |
|
> 60 (Adultos) |
181 |
106,65 ± 33,52 61 - 206 |
76 |
101,76 ± 25,29 61 - 172 |
257 |
101,76 ± 25,29 61 - 172 |
|
Longitud corporal promedio ± D.E. y ámbito de longitud corporal (mm) |
||||||
|
Peso (g) |
n |
Machos |
n |
Hembras |
n |
Ambos |
|
≤ 30 (Jóvenes) |
36 |
93,25 ± 9,13 71 - 112 |
29 |
94,45 ± 10,37 75 - 114 |
65 |
93,78 ± 9,64 71 - 114 |
|
31-60 (Subadultos) |
43 |
118,56 ± 8,93 96 - 135 |
46 |
118,17 ± 9,65 99 - 135 |
89 |
118,36 ± 9,26 96 - 135 |
|
> 60 (Adultos) |
181 |
157,44 ± 16,77 124 - 216 |
76 |
154,33 ± 12,06 129 - 187 |
257 |
156,52 ± 15,56 124 - 216 |
DISCUSIÓN
La información sobre la biología y la ecología de S. hisutus es limitada a unos pocos estudios de corta duración, o bien que incluye a varias especies con poco detalle para cada una, dada la cantidad de especímenes examinados. Si bien esa información es limitada, ha sido útil para disponer de un conocimiento general de una especie que hasta hace relativamente poco era considerada como una subespecie de S. hispidus. La información obtenida en este estudio nos permite disponer de un conocimiento más preciso de esta especie y considerar que la rata de campo (S. hirsutus) es de tamaño mediano con una longitud corporal y peso promedios de 141,65mm y 84,13g y de 131,81mm y 69,07g, para machos y hembras, respectivamente. Esos valores de peso promedio son inferiores a los observados por Monge (2008), ya que para los machos fue de 111,4g (n= 22) y para las hembras de 95,2g (n= 16), para un período de junio a enero, aunque los ámbitos de pesos considerando ambos sexos fueron similares, de 20-206g. Por su parte, de los datos de Vivas & Calero (1985) en un estudio de junio a febrero con muestras de 14 machos y 17 hembras, se deduce un peso promedio superior de los machos de 101,72g y similar para las hembras de 71,89g. Para algunas antiguas subespecies de S. hispidus (S. h. chiriquensis, S. h. borucae y S. h. zanjonensis) que por su distribución geográfica ahora corresponden a S. hirsutus, se reportan valores de 166,5, 152,9 y 140mm, respectivamente (Cameron & McClure, 1988). Para ambas variables (peso y longitud) se determinó que existe un dimorfismo sexual con un tamaño mayor de los machos, similar a lo observado para S. hispidus (Chipman, 1965; Cameron & McClure, 1988).
Con las variables de peso y longitud corporal se estableció una relación que permite estimar cada variable con el valor de la otra, lo cual es útil cuando se dispone de datos de solo una de ellas. En estudios con especímenes vivos, la obtención de peso resulta más fácil, dado que no se requiere de una manipulación excesiva del individuo, y por ende el dato obtenido puede ser confiable, lo cual no siempre ocurre con mediciones de la longitud corporal. En cuanto al uso de un modelo para inferir el valor de una variable en función de otra conocida, debe considerarse que cada modelo se ajusta a las condiciones de los individuos que conformaron la muestra de estudio. La condición corporal de los individuos puede ser afectado por aspectos densodependientes de la población, como fue observado por Odum (1955) en Sigmodon hispidus, cuyo tamaño promedio tendió a incrementarse en periodos de baja densidad.
El índice de condición corporal, estimado de la relación entre la longitud y el peso corporal, osciló entre 1 y 6,4mm/g para los machos y entre 1 y 8,3 mm/g para las hembras. Vivas y Calero (1985) utilizaron esta relación para S. hispidus (ahora S. hirsutus por su distribución geográfica) donde los jóvenes mostraron valores más altos que los adultos, similar a lo observado en este estudio. Los valores menores de este índice indican una mejor condición (Heikura, 1977), es decir mayor peso por unidad de longitud. Al considerar este índice por categoría de peso se obtuvo una relación inversa, lo cual puede ser explicado por un incremento diferencial de las variables peso y longitud a través del tiempo, producto del aumento de peso más que del tamaño conforme aumenta la categoría de peso, la cual puede relacionarse con el aumento de la edad.
La proporción de sexos mostró una mayor representación de machos, similar a lo observado por González, Sandoval, Serna y Aguilar (1978) de 1:0,76 (172 machos y 124 hembras), mientras que Fleming (1970) obtuvo una relación de 1:0,69 (35 machos y 24 hembras) y Monge (2008) de 1:0,72 (22 machos y 16 hembras) sin que se evidenciaran en estos últimos estudios una diferencia significativa de la relación de igualdad, coincidiendo con lo observado por Vivas y Calero (1985) en una relación de 1:1,33 (6 machos y 8 hembras). En este estudio, para las categorías de menor peso (≤ 30 y 31-60g) tampoco se evidenciaron diferencias significativas con la relación de igualdad. La diferencia de proporción de sexos en las categorías de peso superiores a 60 g puede deberse a una mayor mortalidad de hembras durante las primeras categorías de peso o bien a un comportamiento que reduzca su acceso a las trampas y por ende no forman parte de la muestra. En cuanto a la figura que representa la estructura poblacional no sigue la forma piramidal conocida para otras poblaciones, en la que se muestra que las categorías de peso inferior o de menor edad superan a las siguientes, lo cual se explica por la mortalidad que va ocurriendo en cada categoría y por ende las sucesivas van siendo menos representadas. Sin embargo, ese comportamiento es el esperable cuando se le da seguimiento a una cohorte, lo cual no corresponde a este caso.
La actividad reproductiva es poco conocida para esa especie con referencias muy generales, tales como que aparentemente se reproducen todo el año (Vivas & Calero, 1985) o producen jóvenes todo el año (Fleming, 1970), o bien de reportes de reproducción en períodos menores del año (Monge, 2008). En este estudio se observó hembras con inicio o en evidente estado de preñez en todos los meses del año, con una mayor cantidad de capturas en los meses de abril, junio y agosto, lo cual contrasta con lo observado por Fleming (1970) quien no obtuvo capturas de hembras preñadas en los meses de abril y junio. En cuanto a la definición de una época de mayor presencia de hembras preñadas durante el año, se encontró una proporción de hembras preñadas igual o superior al 50% del total de hembras con subadultas y adultas en los meses de abril, agosto, setiembre, diciembre y marzo, pero dado que entre esos meses se encuentran otros con proporciones de hembras preñadas menores, no se puede definir una época de particular actividad reproductiva.
En cuanto al tamaño de camada se obtuvo un promedio de 4,4 (1-7) embriones, siendo menores para las hembras de categorías de peso inferiores (31-60 y 61-90g), lo cual puede estar relacionado con una primera reproducción. Este valor es inferior al obtenido por Monge (2008) de 6,25 embriones. En otros estudios referidos para S. hispidus, pero que por la ubicación de los sitios de estudio dentro del ámbito actual de S. hisrutus, corresponden a esta especie, se obtuvieron tamaños de camada de 2,8 y 5,7 embriones (Bowdre, 1971).
La posición de los testículos de los machos no necesariamente es un parámetro adecuado para determinar la actividad reproductiva de los individuos; sin embargo, la ubicación escrotal de testículos sugiere una condición más propicia para la reproducción que la posición semiescrotal con desarrollo incompleto de escroto y más aún abdominal sin presencia de escroto. En este estudio, durante todos los meses se logró la captura de individuos con testículos escrotales, presentándose una oscilación de cantidad de individuos con testículos escrotales, siendo los meses de mayo (20 individuos), octubre (18), abril de 2009 (13) y setiembre y diciembre (12) con mayor presencia de machos con esta posición testicular, que más que identificar un período específico, sugiere una condición generalizada para un periodo amplio.
Los individuos jóvenes estuvieron presentes en la mayoría de los meses del año, excepto en octubre y enero. El mes con mayor presencia de jóvenes fue abril (12 individuos), seguido de junio (11) y agosto (7), lo que coincide con los meses de mayor presencia de hembras preñadas. La información obtenida de las hembras preñadas, machos con testículos escrotales, así como presencia de juveniles permite deducir que la especie tiene capacidad de reproducirse todo el año, lo que confirma lo sugerido por Fleming (1971) y Vivas y Calero (1985).
Al considerar el tamaño, peso, las características de pelaje, el escaso desarrollo del sistema reproductivo se deduce que los individuos jóvenes pertenecen a la primera categoría de peso (≤30g), mientras que la siguiente categoría (31-60g) fue considerada como la de subadultos, dada las muestras de inicio de preñez en algunos individuos con pesos entre 51 y 58g, similar a lo observado por Goertz (1965) para S. hispidus, cuyas hembras preñadas con menor peso eran de 51 y 53g. En el caso de los machos, en esta categoría de peso (31-60g) se presenta el inicio de desarrollo del escroto. Por su parte, el mayor desarrollo del sistema reproductivo de individuos de la siguiente categoría de peso (61-90g) con mayor proporción de hembras preñadas y machos con desarrollo escrotal, permite asumir que a partir de esta categoría de peso los individuos pueden considerarse adultos. Dada la relación de peso y longitud corporal, podría clasificarse como juveniles a los individuos con peso igual o inferior a 30g y longitud igual o inferior a 105,7mm y la de subadultos de 31-60g y de 105,8 a 134,5mm.
Al comparar estas categorías de peso y su respectiva relación con la edad y desarrollo reproductivo con otras propuestas, tales como la de Odum (1955) para S. hispidus y Monge (2008) para S. hirsutus, se tiene como coincidencia que la primera categoría desde el nacimiento hasta que alcanzan los 30g, conformado por individuos jóvenes sin capacidad para reproducirse. La segunda categoría de peso (31-60g), considerada es este estudio, como constituida por subadultos, corresponde a la segunda parte de la categoría de jóvenes propuesta por Odum (1955) y similar a la de Monge (2008). En la siguiente categoría (61-90g) se cuenta con evidencia de reproducción en aproximadamente la tercera parte de las hembras que conforman esta categoría, la cual corresponde con la primera parte de la categoría llamada adultos jóvenes en las otras clasificaciones. A su vez, la segunda parte de esa clasificación corresponde con la categoría de peso de 90-120g, en la cual hay una evidente actividad reproductiva del 75% de las hembras de la clase. A partir de 120g de peso, los individuos forman parte de la categoría denominada adultos viejos y por ende con una importante actividad reproductiva, que en nuestro estudio oscila entre 83 y 100% de las hembras de esas categorías. Este comportamiento de aumento de la proporción de hembras preñadas con respecto a la cantidad de hembras de la clase, coincide con lo observado por Odum (1955) para S. hispidus.
La especie S. hispidus muestra similitud a S. hirsutus, de ahí que, por mucho tiempo, S. hirsutus se consideró una subespecie de S. hispidus. Esta similitud se refleja en algunas características corporales, tales como el pelaje, así como en sus dimensiones corporales. En cuanto a lo longitud, Jiménez (1971) reportó un tamaño máximo de 217mm para S. hispidus, mientras que para S. hirsutus fue de 216mm, y se ha reportado un peso promedio fue de 88,4g para S. hispidus (Cameron & Spencer, 1988) y para S. hirsutus resultó ser de 84,14 ± 44,30 para los machos y de 69,07 ± 38,85 para las hembras. En este sentido, asumiendo un crecimiento similar entre ambas especies, podría recurrirse a medidas corporales de S. hispidus de edades conocidas, para deducir la edad aproximada de individuos de S. hirsutus a diferentes tamaños. Así, con base en los datos de la relación de peso y edad de Meyer y Meyer (1944) para S. hispidus, los individuos alcanzarían el peso de 30g a los 16 días y los 60g a los 31 y 34 días, para machos y hembras respectivamente. A su vez, según la relación de longitud y edad determinada por Jiménez (1971) también para S. hispidus, el tamaño de 105,7 lo alcanzan a los 17 días, mientras que el de 134,5 mm a los 34 días. Por lo anterior, la edad de joven abarcaría desde el nacimiento hasta los 16-17 días, y la de subadulto de 17 a 31 días para los machos, y de 17-34 para las hembras. Esta diferencia se debe a dimorfismo sexual observado. A partir de los 32 y 35 días se inicia la edad de adulto para machos y hembras, respectivamente. Este límite de 30 g para clasificar a los individuos como juveniles o subadultos coincide con lo indicado por Goertz (1965) para S. hispidus que pierden la apariencia de juveniles en un ámbito de peso de 21 y 40g.
La actividad reproductiva de S. hirsutus mostró una tendencia creciente en la proporción de hembras preñadas con respecto a las hembras a partir de la categoría de peso de 61-90g, la cual fue definida como la de inicio de la edad de adultas. Esto coincide con lo referido por Meyer y Meyer (1944) de que las hembras de S. hispidus inician la reproducción entre los 40 y 50 días, periodo en que tienen un peso entre 62-87 g y guarda relación con lo asumido por Odum (1955) para S. hispidus y Monge (2008) para S. hirsutus, con el inicio de la categoría de jóvenes.
La información obtenida ha permitido ampliar el conocimiento acerca de la biología de la rata de campo (S. hirsutus), lo cual su validez debe corroborarse para otras localidades dentro de su ámbito de distribución. A su vez, considerando la dinámica poblacional de la especie en ciclos de aproximadamente 5 años (Hilje, 1992) conviene considerar el comportamiento de la especie en diferentes momentos del dicho ciclo. De igual manera, es posible que los cambios climáticos que se están presentando puedan inducir a modificaciones en algunos de los aspectos estudiados, mientras que otros por su naturaleza podrían mantenerse según lo obtenido en este estudio.
AGRADECIMIENTOS
El autor agradece a Freddy Osegueda por su colaboración en la toma de datos de campo, así como al apoyo logístico de Jessica Linares y Jorge Álvarez, así como a los productores de Finca Alemania (Santa Cecilia de La Cruz) por facilitar sus áreas de producción para la realización de esta investigación. El estudio fue financiado por la Fundación Proagroin y se contó con el apoyo logístico de la Vicerrectoría de la Universidad de Costa Rica, a través del proyecto 813-A8-514.
REFERENCIAS
Bowdre, L. P. (1971). Litter size in Sigmodon hispidus. The Southwestern Naturalist, 16(1), 126-128. DOI: 10.2307/3670111
Cameron, G. N., & McClure, P. A. (1988). Geographic variation in life history traits of the hispid cotton rat (Sigmodon hispidus). En: Boyce, M. S. (Ed.) Evolution of life histories of mammals. Theory and pattern. (pp. 33-64). Connecticut, USA: Yale University Press.
Carroll, D. S., & Bradley, R. D. (2005). Systematics of the genus Sigmodon: DNA sequences from betafibrinogen and cytochrome b. The Southwestern Naturalist, 50(3), 342-349. DOI: 10.1894/0038-4909(2005)050[0342:SOTGSD]2.0.CO;2
Chipman, R. K. (1965). Age determination of the cotton rat (Sigmodon hispidus). Tulane Studies in Zoology, 12(2), 19-38.
Fleming, T. H. (1970). Notes on the rodent faunas of two Panamanian forest. Journal of Mammalogy, 51(3), 473-490. DOI: 10.2307/1378388
Goertz, J. W. (1965). Sex, age and weight variation in cotton rats. Journal of Mammalogy, 46(3), 471-477. DOI: 10.2307/1377636
González, R. A., Sandoval, T., Serna S. J., & Aguilar R., V. M. (1978). Damages caused by cotton rat, Sigmodon hispidus zanjonensis, on sugar cane in San Pedro Sula, Honduras. Howard, W.E. (Ed.) Proceedings of the 8th Vertebrate Pest Conference. University of Nebraska – Lincoln, Sacramento, California, USA.
Heikura, K. (1977). Effects of climatic factors on the field vole Microtus agrestis. Oikos, 29(3), 607-615. DOI: 10.2307/3543599
Hilje, L. (1992). Biología y ecología de los roedores plaga en Costa Rica. Manejo Integrado de Plagas (Costa Rica), 23,17-25.
Jiménez, J. J. (1971). Comparative post-natal growth in five species of the genus Sigmodon. I. External morphological character relationships. Revista Biología Tropical, 19(1,2), 133-148.
Krebs, C. J., & Singleton, G. R. (1985). Indices of condition for small mammals. Australian Journal of Zoology, 41, 317-23. DOI: 10.1071/ZO9930317
Layne, J. N. (1974). Ecology of small mammals in a flatwoods habitat in north-central Florida, with emphasis on the cotton rat (Sigmodon hispidus). American Museum Novitates, 2544, 1-48.
Meyer, B. J., & Meyer, R. K. (1944). Growth and reproduction of the cotton rat, Sigmodon hispidus hispidus, under laboratory conditions. Journal of Mammalogy, 25(2), 107-129. DOI: 10.2307/1375009
Monge, J. (2008). Estructura poblacional y actividad reproductiva de la rata de campo (Sigmodon hirsutus) durante un ciclo de producción de maní (Arachis hypogaea) en Costa Rica. Agronomía Costarricense, 32(2), 161-167.
Odum, E. P. (1955). An eleven year history of a Sigmodon population. Journal of Mammalogy, 36(3), 368-378. 10.2307/1375679 DOI:
Peppers, L. L., Carroll, D. S., & Bradley, R. A. (2002). Molecular systematics of the genus Sigmodon (Rodentia:Muridae): evidence from the mitochondrial cytochrome-b gene. Journal of Mammalogy, 83(2), 396-407. DOI: 10.1644/1545-1542(2002)083<0396:MSOTGS>2.0.CO;2
Vivas, A. M., & Calero, A. C. (1985). Algunos aspectos de la ecología poblacional de los pequeños mamíferos en la estación Biológica de Los Llanos. Boletín de la Sociedad Venezolana de Ciencias Naturales, 143,79-99.