Estructura poblacional de ocho especies maderables amenazadas
en el departamento del Chocó-Colombia
Luz Yorleyda Palacios Tello1, Keiler Perea Pandales2, Danilza Marcela Bellido1, Haylin Yineth Caicedo3 Moreno & Deivis Abadia Bonilla1
1. Grupo de investigación de Flora Chocoana, Programa de Biología, Chocó, Colombia, Facultad de Ciencias Naturales, Universidad Tecnológica del Chocó “Diego Luis Córdoba”; yorleida16@hotmail.com, damabeco02@gmail.com, deivisabadia@gmail.com
2. Grupo de investigación en recursos vegetales, Programa de Biología, Chocó, Colombia, Facultad de Ciencias Naturales, Universidad Tecnológica del Chocó “Diego Luis Córdoba”; keilerpereapandales@yahoo.es
3. Grupo de investigación de Ecología y Conservación de Ecosistemas Tropicales, Programa de Biología, Chocó, Colombia, Facultad de Ciencias Naturales, Universidad Tecnológica del Chocó “Diego Luis Córdoba”; haylincaicedo@gmail.com
Recibido 08-viii-2016 • Corregido 16-X-2016 • Aceptado 27-i-2017
Abstract: Population structure of eight threatened timber species in the Department of Chocó-Colombia. This study was conducted in order to evaluate the population structure and the current status of seven timber species for Chocó, where a plot of 50x50, which were recorded in a zig zag all individuals with DBH≥10cm was established (adults). From the mother tree they were measured 15m radius (707m2 ) where a circular plot in which all individuals over 1.3m high and lower were recorded at 10 cm of DAP ( juveniles) was installed ; following the diagonals of the first plot 12 plots of 2m (4m2 ) where all individuals were recorded height between 0.3 and 1.30m were drawn . At the species level were recorded at Hymenaea oblogifolia and Dipterix oleifera with the highest density values , a total density of 19.8 ind/ha was found. Most individuals were located in the diametric class 30.1 to 40; height class for a form of inverted J secured the sustainability of the species was found.
Keywords: community, Population density, distribution, successional stage, ecology, uniformity.
El nivel de integración de la materia que denominamos población, tiene una posición intermedia entre los individuos y las comunidades ecológicas. Debido a esta posición, y a las íntimas relaciones que tiene con los niveles inferiores y superiores inmediatos, los estudios de distribución y abundancia de las poblaciones son una parte esencial de la ecología (Rumi, 2012). El estudio de la distribución espacial de las especies es un factor fundamental para comprender o determinar el efecto de sucesos pasados sobre el patrón de distribución actual, generando hipótesis sobre los procesos biológicos o ambientales que estructuran o interviene en los bosques tropicales (Montañez et al., 2010). Una de las características más conspicuas de la biodiversidad es que no se distribuye de manera homogénea, por lo que los patrones espaciales de la diversidad de especies y los procesos asociados a los mismos, han sido objeto de estudio desde hace mucho tiempo y desde diferentes escalas y enfoques como son ecología de comunidades, la biogeografía, y la macro ecología (Koleff et al., 2008).
Diversos autores señalan a los bosques húmedos tropicales como ecosistemas de gran importancia por sus altos valores de biodiversidad (citar Gentry Primack et al., 2001a; Hartshorn, 2002; Brooks et al., 2002), de la que los humanos han hecho uso para satisfacer sus necesidades y han sido importantes para su sobrevivencia pasada, presente y futura (Weaver y Bauer, 2000; Wadsworth, 2000; Primack et al, 2001b).
Vale la pena mencionar algunos autores que han realizado investigaciones abordando los temas de densidad poblacional y distribución espacial, con el propósito de contribuir al conocimiento de la diversidad forestal.
Para ello reviste la importancia de Beltrán K et al. (2009), Cabrera y Wallace (2007), Dávila et al. (2012), Parrado (2007), Guerrero et al. (2010), Montañez et al. (2010), Vallejo et al. (2009), Koleff et al. (2008), Arellano y Rangel (2008).
En el departamento del Chocó algunos autores contribuyen al conocimiento de densidad poblacional y distribución espacial como lo muestran las investigaciones de Palma y Andrades (2010), Andrade y Murillo (2012), Palacios et al. (2015), Ramírez et al. (2009), Robledo-Murillo (2009).
La región del Chocó es considerada como una de las regiones de mayor biodiversidad a nivel mundial, presenta varios tipos de ecosistemas y un alto endemismo tanto de especies florística como faunística lo que hace nombrarlo “Chocó biogeográfico” (Brown, 1982), Gentry, (1986) revela que este endemismo se debe al aislamiento de la región del resto de las demás tierras bajas de Suramérica por la cordillera de los Andes, por ende posee bosques húmedos Tropicales (bh-T) y bosques muy húmedos Tropicales (bmh-T). En este sentido, la presente investigación tuvo como propósito, determinar la estructura poblacional y el estado actual de 8 especies forestales para chocó. Como un aporte al conocimiento del estado de las especies maderables.
MATERIALES Y MÉTODOS
Área de estudio: El Chocó es uno de los treinta y dos departamentos de Colombia, localizado en la región del Pacífico en el noroeste del país entre los 04º00’50’’ y 08º41’32’’ de latitud Norte y los 76º02’57’’ y 77º53’38’’ de longitud Oeste. Comprende las selvas del Darién y las cuencas de los ríos Atrato, San Juan y Baudó. Siendo su capital la ciudad de Quibdó; es este es considerado el lugar con la mayor pluviosidad del planeta. Tiene una extensión de 46.530 Km2 (11.497.813 acres), que corresponde al 4% de la extensión del país.
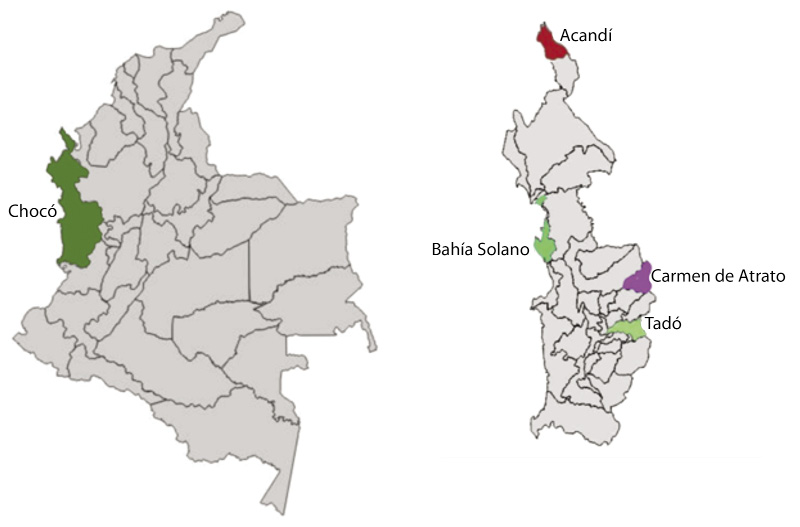
Descripción de los sitios de estudio: Este estudio se realizó en cuatro municipios, pertenecientes al departamento del Chocó, Acandi está situado a los 8°30’41” N 77°16’40” en el extremo norte del Chocó, noroccidente de Colombia, a orillas del mar caribe; Bahía Solano se localiza a los 6°13’27” N 77°24’14” O, en la región de las calmas ecuatoriales. Carmen de Atrato, el cual se encuentra ubicado al occidente del departamento a los 5º 54’40’’ N, 76º12’30’’ W; Tado se ubica a los 5º 16’ 10” de latitud norte de la línea del Ecuador y a los 76º 73’ 10” de longitud oeste del meridiano de Greenwich (zona centro-oriente del departamento del Chocó) (Fig. 1).
Para el cumplimiento del objetivo en campo se utilizó la metodología propuesta por Lamprecht (1990) con algunas modificaciones, para cada especie se estableció una parcela de 50x50 (2500m2 equivalente 0,25 ha), donde se registraron en forma de zig zag todos los individuos con DAP ≥ 10 cm (adultos). A partir del árbol madre se midieron 15m de radio (707m2) donde se estableció una parcela circular en la cual se registraron todos los individuos mayores a 1.3 m de altura y con DAP ≤ 10 cm de DAP (juveniles); siguiendo las diagonales de la primera parcela se trazaron 12 cuadriculas de 2m (4m2) donde se censaron todos los individuos con altura entre 0.3 y 1.3 m (plántulas) (Fig. 2)
Índice de Morisita
I∂ = N((∑ni (ni - 1))/(n (n - 1)))
Dónde:
I∂ = índice de Morisita.
N = número de muestras.
ni = número de individuos en la i-ésima muestra.
n = número total de indivíduos en todas las muestras.
Densidad: Los valores correspondientes al número de individuos para cada una de las especies fueron utilizados para determinar el tamaño de sus poblaciones. Esta se calculó mediante el conteo simple del número total de individuos por área muestreada.
Categoría de Tamaños de la Especies: Los datos obtenidos (densidad, alturas, DAP) de las siete especies forestales se utilizaron para determinar la distribución de las categorías de clase diametrica y de altura de las mismas.
RESULTADOS
Composición florística: Se registraron 595 individuospertenecientes a 5 familias, 8 géneros y 8 especies.
Las especies evaluadas fueron Anacardium excelsum, Carapa guianensis, Cedrela odorata, Dipterix oleífera, Huberodendron patinoi, Humiriastrum procerum, Hymenaea oblongifolia, Terminalia oblonga. Las familias más abundantes fabaceae y Anacardiaceae con 293 y 135 individuos respectivamente (Cuadro 1).
A nivel de especies las másabundantes son Hymenaea oblogifolia, Dipterix oleíferacon 147 y 146 individuos respectivamente, la menos abundante fue Huberodendron patinoi con 10 individuos.
En cuanto a la regeneración ninguna de las especies estuvo presente en todas los estados sucesionales de crecimiento, la especie H. patinoi solo presento individuos adultos, H. oblongifolia y D. oleífera sobresalen por presentar la mayor abundancia de individuos en estado plántula.
Densidad Poblacional: La densidad total fue de 19,8 ind/ha, de los cuales las plántulas registraron 0,069 ind/m2, los juveniles 0,019 ind/m2 y los adultos 6,01ind/m2.Las plántulas estuvieron mejor representadas con 400 individuos, seguido de los adultos con 181 individuos.
A nivel de especies sobresalen D. oleífera (58,4 ind/ha) H. oblogifolia (58,8 ind/ha) y A. excelsum (54 ind/ha). Cuadro 2.
Los municipios más abundantes fuero Acandí (19,2 ind/ha). Bahía solano (17,3 ind/ha) y Tadò (31,4/ha), mientras que el Carmen de Atrato (14,6 ind/ha) fue el que menos individuos registro
La prueba Anova mostro que existen diferencias estadísticamente significativas entre los estados sucesionales plántulas-Latizal, y plántula-fustal (Fig. 3). En cuanto a la densidad poblacional entre municipios no existe diferencias estadísticamente significativa entre ellos (Fig. 4).
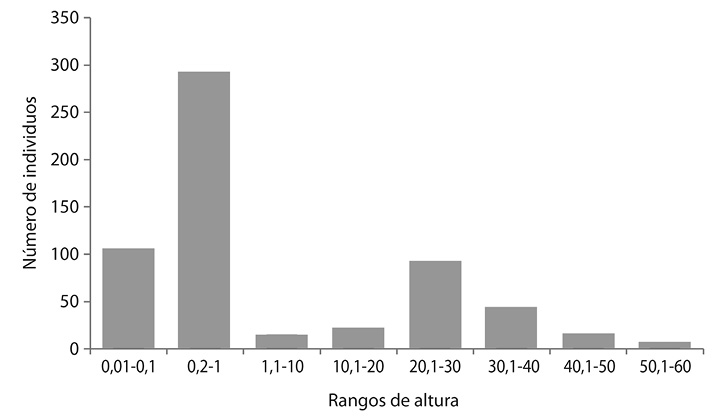
Distribución de altura: En cuanto a la distribución de altura La mayor cantidad de individuos se ubicaron en los rangos de 0,01-0,1 y 0,2-1, mostrando diferencias significativas en los distintos rangos (Fig. 5).
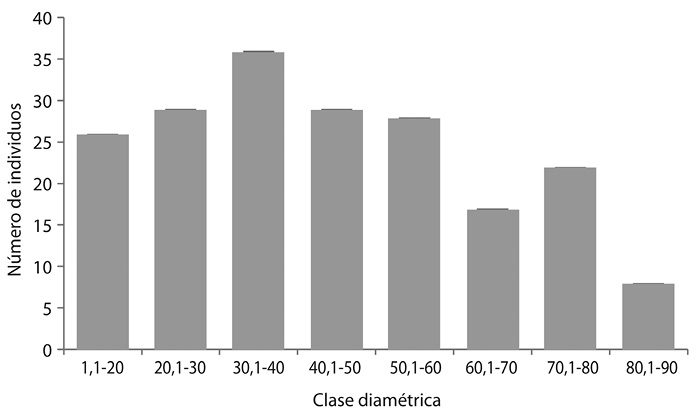
Para las clases diamétrica se encontró que la mayor cantidad de individuos se ubicó en el rango de 30,1-40, mostrado una estabilidad en los rangos intermedios, mientras que el Rango de 80,1-90 presento una disminución significativa en el número de individuos (Fig. 6).
Patrón de distribución espacial: El índice de dispersión de Morisita en las especies de Cedrela odorata (0,1), Terminalia oblonga (0,14), Humiriastrum procerum (0,067), Dipterix oleífera (0,059) Hymenaea oblongifolia, (0,607)Carapa guianensis (0,02) Huberodendron patinoi (0,025), en las cuales se evidencia una distribución uniforme y la especie Anacardium excelsum (1,02) es la única especie con una distribución agregada (Cuadro 3).
DISCUSIÓN
Las estructuras poblacionales son un componente importante de la evaluación de las especies (Gallego & Finegan, 2004). El análisis de las distribuciones diamétrica nos permite inferir el estado demográfico de las masas de bosque y sus posibles problemas de conservación, (R. Ajbilou et al., 2003). En este trabajo se encontró una distribución diamétrica en forma de montículo, lo que según Moreno (2004), se debe a la proporción de abundancia de plántulas, es decir que bosques con alta densidad de plántulas presenta una población grande, y bosques con pocas plántulas presentan una población significativamente pequeña, mientras Finegan (1996), plantea que esta distribución en forma de montículo puede deberse al crecimiento rápido de algunos árboles y la mortalidad de los menos favorecidos.
En cuanto a la distribución de altura, Nascimiento et al. (1997) afirma que la distribución en forma de j invertida en el bosque secundario refleja un reclutamiento regular de nuevos individuos en la población; lo que es predecible para una especie secundaria que se desarrolla en áreas abiertas. Lo anterior concuerda con Araujo-Murakami (2002), quien argumenta que, esta disminución de individuos se explica a través de la dinámica de crecimiento y mortalidad, los individuos muertos y los ascendidos a la clase inmediata superior son reemplazados por aquellos reclutados de la clase inmediata inferior, los cuales mantienen la estructura del bosque.
El estudio de la distribución espacial en los ecosistemas forestales es de gran interés en lo correspondiente al manejo sostenible de los recursos naturales. Por un lado está relacionada con el crecimiento de los árboles y por tanto, de las masas arboladas. La inclusión en la toma de datos de la posición de los individuos en los inventarios forestales ha servido como punto de partida para las investigaciones sobre las distribuciones espaciales de las masas forestales (Füldner et al., 1994). Algunas poblaciones se encuentra agregadas en los sitios donde han ocurrido claros producidos por la caída de árboles, otras se agrega a pobres niveles de dispersión de semillas y baja depredación de las mismas, y otras muestra agregación en relación con hábitats topográficos y edáficos específicos (Martiez-Ramos y Alvarez-Builla, 1995).Teniendo en cuenta lo anterior y observando las características topográficas, edáficas y los claros producidos por perturbaciones naturales y antrópicas, se puede dar una posible respuestas a que la especie Anacardium excelsum presente un patrón de distribución agregada ya que esta especie permanece en bosques inundables o cerca de los ríos con una alta disponibilidad de luz.
Mientras que el resto de las especies objeto de estudio presentaron un patrones de distribución uniforme, esto puede deberse según Morlans (2004), únicamente a factores intrínsecos de las especies, dado que es difícil suponer que de manera natural los recursos se dispongan equidistantes en el espacio, la competencia o el comportamiento agresivo intraespecífico parecen ser los principales agentes causales de un patrón uniforme, ya que es en este patrón en el cual la supervivencia se maximiza y las interacciones hostiles se llevan a un mínimo.
Los municipios realizó donde se estudió fueron Acandí, Bahía, Tadò, Carmen de Atrato Se registrándose 8 especies, siendo Hymenaea oblogifolia con 147 individuos. Las familias mejor representadas fueron Meliaceae y Fabaceae con 2 individuos respectivamente. En cuanto a la densidad poblacional se encontró un densidad total de 19.4 ind/ha.
AGRADECIMIENTOS
Esta investigación fue financiada con recursos de regalías (Gobernación del Chocó) y ejecutado por la Universidad Tecnológica del Chocó “Diego Luis Córdoba” en el marco del proyecto “Aplicación de la CTeI para el Mejoramiento del sector Maderero en el departamento del Chocó”. Los investigadores del proyecto Agradecemos la hospitalidad y colaboración de los habitantes de los municipios del Carmen de Atrato (guaduas), Bahía Solano (comunidad indígena de boro boro), Tadó (angostura) y Acandi (Acandi seco).
REFERENCIAS
Ajbilou, R., Marañón, T., & Arroyo, J. (2003). Distribución de clases diamétricas y conservación de bosquesen el norte de Marruecos. Invest. Agrar.: Sist. Recur. For. 12(2), 111-123.
Andrade, E., & Murillo, A. S. (2012). Densidad poblacional de Hymenaea oblongifolia Huber. En el municipio de Quibdó, chocó – Colombia. Trabajo de grado para obtener el título de Biólogo con énfasis en recursos naturales. Tesis de grado. Quibdó-Chocó. Universidad Tecnológica del Chocó.
Araujo-Murakami, A. (2002). Dinámica, incorporación y almacenamiento de biomasa y carbono en el Parque Nacional Noel Kempff Mercado. Tesis de Licenciatura en Ingeniería Forestal, Universidad Autónoma Gabriel René Moreno, Santa Cruz, Bolivia.
Arellano, H., & Rangel, J. O. (2008). Patrones en la distribución de la vegetación en áreas de páramo de Colombia: heterogeneidad y dependencia espacial. Revista Caldasia 30(2), 355-411.
Barreto-Silva, J. S., Cárdenas, D., & Duque Montoya, Á. J. (2013). Patrones de distribución de especies arbóreas de dosel y sotobosque a escala local en bosques de tierra firme, Amazonia colombiana. Revista Biología Tropical, 62(1), 373-383 pp.
Beltrán, K., Salgado, S., Cuesta, F., León-Yánez, S., Romoleroux, K., Ortiz, E., Cárdenas, A., & Velástegui, A. (2009). Distribución Espacial,Sistemas Ecológicos y Caracterización Florística de los Páramos en el Ecuador. EcoCiencia, Proyecto Páramo Andino y Herbario QCA. Quito.
Brooks, T., Mittermeier, R., Mittermeier, C., Da Fonseca, G., Rylands, A., Konstant, W., Flick, P., Pilgrim, J., Oldfield, S., Magin, G., & Hilton-Taylor, C. (2002). Habitat loss and extinction in the Hotspots of biodiversity. Conservation Biology, 16, 909-923.
Cabrera, W. H., & Wallace. R. (2007). Densidad y distribución espacial de palmeras arborescentes en un bosque preandino-amazónico de Bolivia. Revista Ecologia en Bolivia, 42 (2), 121-135.
Dávila, D. E., Alvis, S. F., & Ospina, R. (2012). Distribución espacial, estructura y volumen de los bosques de roble negro (colombobalanus excelsa (lozano, hern. cam. & henao, j.e.) nixon & crepet) en el parque nacional natural cueva de los guácharos. Revista Colombia Forestal, 15(2), 207-214 pp.
Finegan, B. (1996). Pattern and process in neotropical secondary rain forest: the first hundred years of succession. Trends in Ecology and Evolution, 11, 119-124.
Füldner, K., & Gadow, K. (1994). How to define a thinning in a mixed deciduo beech forest. In: Mixed Stands. Proceeding from the symposium of the IUFRO Working Groups S4. 01-03 and S4. 01-04. Lousa & Coimbra, Portugal, 31-42.
Gallego, B., & Finegan, B., (2004). Evaluación de enfoques para la definición de especies arbóreas indicadoras para el monitoreo de la biodiversidad en un paisaje fragmentado del corredor biológico mesoamericano. Recursos Naturales y Ambiente, 41, 49-61.
Gentry, A. (1982). Phytogeographic patterns as evidencefor a Chocó refuge. En: Biological Diversification In The Tropics. G. Prance (Ed.). Capítulo X, Págs. 112-136; Columbia University.
Guerrero-Rodríguez, A. B., Paz-Camacho, E. A., & Parrado-Rosselli, A. (2010). Efecto de la intervención antrópica en la distribución de las semillas y plántulas del roble (Quercus humboldtii bon pl., fagaceae) en la cordillera oriental colombiana. Revista Colombia Forestal, 13 (1), 163-180 pp.
Hartshorn, G. (2002). Biogeografía de los bosques neo tropicales. In Guariguata, M., Kattan, G. Eds. Ecología y conservación de bosques neotropicales. LUR. Cartago, Costa Rica. P. 59-81.
Koleff, P., & Soberón J. (2008). Patrones de diversidad espacial en grupos selectos de especies, en Capital natural de México, Conocimiento actual de la biodiversidad Vol.1, Conabio, México, pp. 323-364.
Martínez-Ramos, M., & Alvarez-Buylla, E. (1995). Ecología de poblaciones de plantas en selvas húmedas de México. Boletín dela Sociedad Botánica de México, 56, 121-153.
Montañez, R. A., Escudero, C. Y., & Duque, Á. J. (2010). Patrones de Distribución Espacial de Especies Arbóreas en Bosques de Alta Montaña del Departamento de Antioquia, Colombia, Revista Facultad Nacional de Agronomía, 63(2), 5629-5638.
Moreno, S. (2004). Estructura poblacional del roble negro (Colombobalanus excelsa) en el valle del cauca, una herramienta para la formulación de una estrategia para su conservación. Proyecto de grado Biólogo, Universidad ICESI, Facultad de Ciencias Naturales, Biología, Santiago de Cali.
Morláns, M. C. (2004). Introducción a la ecología de poblaciones, Area ecología, científica universitaria, Universidad Nacional de Catamarca, Argentina. 16 pág.
Nascimento, M. T., Proctor, J., & Villela, D. M. (1997). Forest structure, floristic composition and soils of an Amazonian monodominant forest on Maraca’ Island, Roraima, Brazil. Edin J. Bot. 54, 1-38.
Palacios, L. Y., Palacios, J. C., & Abadia, D. (2015). Densidad poblacional de Brosimum utile en un bosque con actividades de minería y tala en el Chocó, Colombia. Cuadernos de Investigación UNED, 7(2), 319-323.
Palma, L. D., & Andrades, S. (2010). Distribución espacial de Cedrela odorata en el municipio de Lloró, Chocó-Colombia. Trabajo de grado para obtener el título de Biólogo con énfasis en recursos naturales. Tesis de grado. Quibdó- Chocó. Universidad Tecnológica del Chocó.
Primack, R., Rozzi, R., Dirzo, R., & Feinsinger, P. (2001a). ¿Dónde se encuentra la diversidad biológica? In Primack, R., Rozzi, R., Feinsinger, P., Dirzo, R., Massardo, F. Eds. Fundamentos de conservación biológica: perspectivas latinoamericanas. Fondo de Cultura Económica. México, D. F. p. 99-129.
Primack, R., Rozzi, R., Dirzo, R., & Feinsinger, P. (2001b). Destrucción y degradación del hábitat. In Primack, R., Rozzi, R., Feinsinger, P., Dirzo, R., Massardo, F. Eds. Fundamentos de conservación biológica: perspectivas latinoamericanas. Fondo de Cultura Económica. México, D. F. p. 183-223.
Ramírez-Moreno, G., & Galeano G., (2009). Comunidades de palmas en dos bosques de Chocó, Colombia. Revista Caldasia, 33 (2), 315-329 pp.
Ramón P. (2009). Modelos en ecología: un enfoque a la distribución espacial de especies vegetales mediante procesos de puntos, Instituto de Ecología, Unidad de Ecología, Universidad Técnica Particular de Loja. Loja, Ecuador.
Robledo-Murillo, D. (2009). Implementación de estudio base para especies forestales amenazadas, en el municipio de Alto Baudó, departamento del Chocó, Colombia, Revista Bioetnia, 6 (2), 82-92 pp.
Vallejo, M. I., & Galeano, G. (2009). Cambios a corto plazo en los patrones de distribución espacial de nueve especies de plantas comunes en un bosque nublado al sur-occidente de Colombia, Revista Caldasia, 31(1), 77-98.
Wadsworth, F. (2000). Producción forestal para América tropical. USDA Forest Servicie. Washington, USA. 603 p.
Weaver, P., & Bauer, G. (2000). Major Meliaceae in Nicaragua. USDA Forest Service. Río Piedras, Puerto Rico.
Resumen: Este estudio se realizó con el objetivo de evaluar la estructura poblacional y el estado actual de 8 especies maderables para el Chocó, donde se estableció una parcela de 50x50, donde se registraron en forma de zig zag todos los individuos con DAP≥10cm (adultos). A partir del árbol madre se midieron 15m de radio (707m2) donde se instaló una parcela circular en la cual se registraron todos los individuos mayores a 1.3 m de altura y menores a 10cm de DAP (jóvenes); siguiendo las diagonales de la primera parcela se trazaron 12 parcelas de 2m (4m2) donde se registraron todos los individuos con altura entre 0.3 y 1.30m. A nivel de especie se registraron a Hymenaea oblogifolia y Dipterix oleífera con los valores más altos de densidad, se encontró una densidad total de 19,8 ind/ha. La mayoría de los individuos se ubicaron en la clase diametrica de 30,1-40; para la clase de altura se encontró una forma de J invertida asegurado la sostenibilidad de las especies.
Palabras clave: comunidad, densidad poblacional, patrones de distribución, clases diamétricas, clases de alturas.
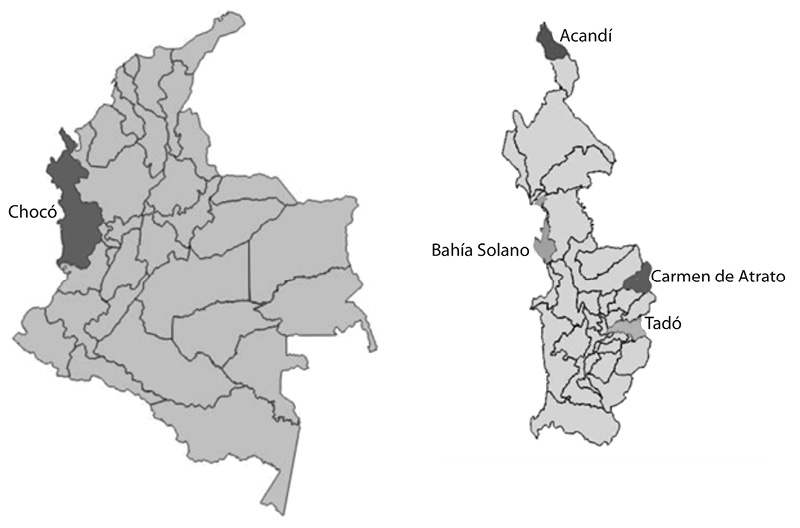
Fig. 1. Localización de los sitios de estudio.
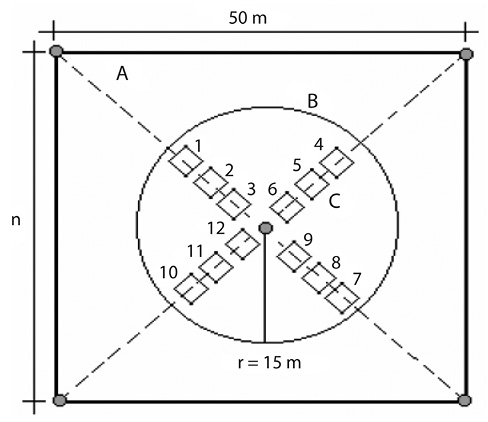
Fig. 2. Metodología de los compartimientos, para la evaluación de la regeneración natural en ecosistemas boscosos tropicales: A= parcela cuadrada de 50 m de lado. B= parcela circular de 15 m de radio. C= conjunto de 12 parcelas cuadradas de 2 m de lado. Fuente: Lamprecht (1990) tomado de Melo y Vargas (2003).
CUADRO 1
Composición de las ocho especies forestales evaluadas en el departamento del Chocó
|
Nombre vulgar |
Nombre científico |
Familia |
Municipio |
|
Chanó |
Humiriastrum procerum |
Humiriaceae |
Bahía solano |
|
Carrá |
Huberodendron patinoi |
Malváceae |
Tadó |
|
Algarrobo |
Hymenaea oblongifolia |
Fabaceae |
Tadó |
|
Choibá |
Dipterix oleífera |
Fabaceae |
Acandí |
|
Cedro |
Cedrela odorata |
Meliaceae |
Acandí, Carmen de Atrato |
|
Caracolí |
Anacardium excelsum |
Anacardiaceae |
Acandí, Bahía solano |
|
Guayabon |
Terminalia oblonga |
Combretaceae |
Carmen de Atrato, Bahía solano |
|
Güino |
Carapa guianensis |
Meliaceae |
Acandí, Bahía solano |
CUADRO 2
Densidad poblacional de los individuos adultos de las especies forestales estudiadas
|
Especie |
Brinzal |
Area muest. ha |
Densidad ind/ha |
Latizal |
Area muest. ha |
Densidad ind/ha |
Fustal |
Area muest. ha |
Densidad ind/ha |
|
Carapa guianensis |
2 |
0,096 |
41,666667 |
0 |
1,414 |
0 |
24 |
5 |
13,6 |
|
Cedrela odorata |
5 |
0,096 |
52,083333 |
14 |
1,414 |
19,8019802 |
24 |
5 |
13,6 |
|
Humiriastrum procerum |
2 |
0,048 |
41,666667 |
0 |
0,707 |
0 |
16 |
2,5 |
6,4 |
|
Hymenaeae oblongifolia |
138 |
0,048 |
2875 |
0 |
0,707 |
0 |
9 |
2,5 |
3,6 |
|
Dipterix oleifera |
136 |
0,048 |
2833,3333 |
0 |
0,707 |
0 |
10 |
2,5 |
4 |
|
Huberodendron patinoi |
0 |
0,048 |
0 |
0 |
0,707 |
0 |
10 |
2,5 |
4 |
|
Anacardium excelsum |
94 |
0,048 |
1958,3333 |
0 |
1,414 |
0 |
43 |
5 |
8,6 |
|
Terminalia oblonga |
25 |
0,048 |
520,83333 |
0 |
1,414 |
0 |
47 |
5 |
9,4 |
Fig. 3. Densidad poblacional entre estados sucesionales de desarrollo de las especies forestales estudiadas. Prueba f = 3,56; P-Valor = 0,0267 existe una diferencia estadísticamente significativa con un nivel del 95,0% de confianza.
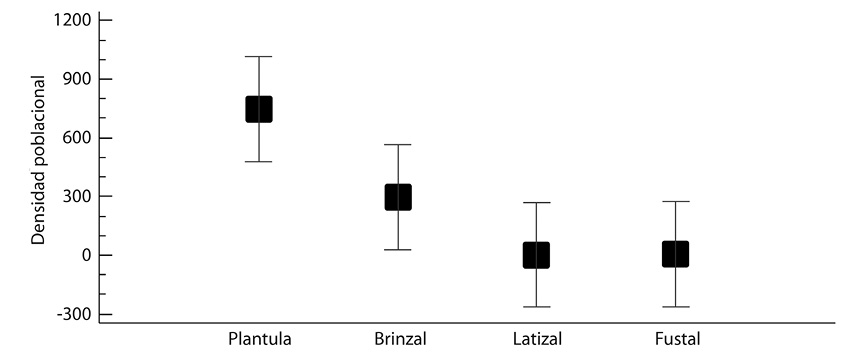
Fig. 4. Densidad poblacional por entre las zonas de estudio. Prueba F= 0,28; Valor-P= 0,8370.
No existe una diferencia estadísticamente significativa con un nivel del 95,0% de confianza.
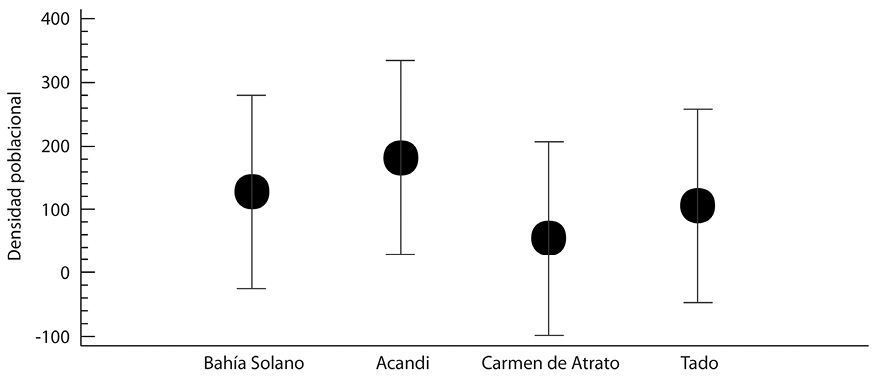
Fig. 5. Distribución por clase de alturas de las siete especies forestales.
CUADRO 3
Distribución espacial de las siete especies forestales
|
Especie |
IN. Morisita |
Tipo de distribución |
|
Cedrela odorata |
0,1 |
Uniforme |
|
Anacardium excelsum |
1,02 |
Agregada |
|
Terminalia oblonga |
0,14 |
Uniforme |
|
Humiriastrum procerum |
0,067 |
Uniforme |
|
Dipterix oleifera |
0,059 |
Uniforme |
|
Hymenaea oblongifolia |
0,607 |
Uniforme |
|
Carapa guianensis |
0,02 |
Uniforme |
|
Huberodendron patinoi |
0,025 |
Uniforme |
Fig. 6. Distribución por clase diametrica de las siete especies forestales.