Riqueza y caracterización ecológica de aves
en bosque nativo y plantaciones exóticas (Prusia, Costa Rica)
Mónica González
Escuela de Ciencias Exactas y Naturales, Universidad Estatal a Distancia(UNED), Mercedes de Montes de Oca, San José, Costa Rica, 474-2050; mgonzalezn@uned.ac.cr
Recibido 02-ii-2017 • Corregido 24-iii-2017 • Aceptado 26-iv-2017
ABSTRACT: Avifaunal species richness in native forest and exotic plantations: effect of altitude and vegetation (Prusia, Costa Rica). The Prusia sector of the Irazú Volcano National Park, Costa Rica, has plantations of native and exotic species, the latter includes pine, cypress and eucalyptus. I estimated the avifaunal richness with point counts (fixed-radius), in pine, eucalyptus and forest, 36 species distributed in 14 families were recorded. Eucalyptus and the forest share 90% of species. The type of habitat and the vertical species distribution were the unique two variables that showed to be associated with the type of vegetation. The low species richness found, in this study, could be explained due to altitude zone and the homogeneity of exotic vegetation patches, that could cause a decrease of resources and shelter sites, while the lack of ecological associations with the type of habitat, could be the result from edge effect, distance from the nature reserve, presence of a middle vegetation layer, or some other factors.
Key words: Richness, birds, Costa Rica, forest, exotic plantations.
El llamado “Sector Prusia” (Parque Nacional Volcán Irazú, Costa Rica) tiene áreas boscosas que fueron arrasadas por la erupción volcánica de 1963-1965. Para estabilizar, proteger el suelo y evitar derrumbes adicionales, se cultivaron rodales de especies nativas como Alnus acuminata (Jaúl) y especies exóticas como el Pinus pseudostrobus, Cupressus sp. y Eucalyptus sp. Hasta ahora no se había estudiado el efecto de esta modificación del bosque natural (Murata, Dondoli & Saénz, 1966; Fonseca, 2009) sobre la avifauna.
Las plantaciones exóticas tienen efectos beneficiosos y perjudiciales sobre los organismos. Pueden tener una función catalizadora en la recuperación de áreas degradadas (Castaño, Morales & Bedoya, 2008) así como jugar una función clave en la conservación de las aves, representando áreas de alimentación, descanso y anidamiento (Castaño & Patiño, 2000, Marsden, Whiffin & Galetti, 2001; Hobbs et al; 2003).
Por otro lado, las plantaciones presentan características ecológicas distintas de los bosques naturales (Hobbs et al., 2003). Esta transformación del ecosistema, genera una disminución de la calidad del hábitat, reflejándose en una escasez de recursos que afecta a la fauna. La capacidad de adaptación es diferente entre organismos; unos pueden ser beneficiados y otros perjudicados ante la modificación del hábitat (Collazo & Bonilla 1988; Castaño & Patiño 2000; Santos & Tellería, 2006; Martínez, 2008).
En este estudio, se estimó la riqueza de aves en tres tipos de vegetación, bosque, pino y eucalipto. Este tipo de investigación proveeerá una base para otras investigaciones y aporta información importante sobre la ecología del sitio, para un uso adecuado de sus recursos (Guariguata & Ostertag, 2001).
METODOLOGÍA
El Sector Prusia (09°95’35”N - 083°88’13” W, Niehaus et al., 2012; Fig. 1) tiene una precipitación promedio de 1527,9mm anuales; sin embargo, es menor a finales de julio y principios de agosto. La estación lluviosa se prolonga durante todo el año, con excepción de febrero y marzo y su temperatura anual es de 14°C (Bermúdez & Conejo, 2008). Este sitio presenta dos zonas de vida, bosque muy húmedo montano y bosque pluvial montano, los cuales se encuentran a una altitud entre 2400 a 3700 m.s.n.m. (Quesada, 2007; Bermúdez & Conejo, 2008).
Para la determinación de la riqueza de la avifauna en el bosque, pinar y eucaliptos, se utilizó la metodología de conteo por puntos con radio definido (Ralph et al., 1996). Los recorridos se realizaron en senderos sin pavimentar, tanto principales como secundarios dentro del sector de Prusia. En los casos en que se utilizaron senderos pavimentados, los conteos se desarrollaron manteniendo una distancia de 3 a 5m de la orilla del sendero (cuando las condiciones del terreno lo permitieron), con el fin de evitar perturbación por turistas (Ralph et al., 1996).
Para evitar efecto de borde dentro de cada ecosistema, se estableció el primer sitio de muestreo a 50m de su límite, a partir del cual, cada 100m, se marcaron los siguientes puntos de conteo. Se muestrearon 64 puntos (31 puntos en bosque, 16 en el pinar y 17 en los eucaliptos). Todos los puntos se ubicaron entre los 2400- 3700 m.s.n.m. (Blake & Loiselle, 2000). En cada punto de conteo se anotó durante 5 minutos, todas las especies observadas y escuchadas dentro de un radio de 25m; este registro se complementó con grabaciones para un análisis posterior. Si en el momento de acercarse al punto de conteo se observó un ave cerca de éste, la especie se anotó sólo si ninguna otra se había detectado durante todo el muestreo (Ralph et al., 1996; Sánchez et al., 2011). Los puntos fueron permanentes y georreferenciados (Ralph et al., 1996; Cerezo, Robbins & Dowell, 2009; Sánchez, 2009; Ramírez, 2010).
Los muestreos se realizaron una vez al mes durante 6 meses, del 22 de octubre del 2010 al 03 de abril del 2011. Para reducir los sesgos por la secuencia de horario (Ugalde et al., 2009), mensualmente se cambió el orden de la secuencia de los sitios. Los recorridos se realizaron durante las mañanas, entre las 5:30 y 9:30am, de esta manera se redujo la variabilidad por condiciones climáticas y pico de actividad de las aves; además de que éste es el período más estable en términos de observación (Blake, 1992; Ralph et al., 1996; Botero, 2005).
Para el arreglo sistemático de las aves, se utilizó la lista oficial de las aves de Costa Rica de la Asociación Ornitológica de Costa Rica (AOCR), ٢٠١٥-٢٠١٦. Para clasificar las especies según el tipo de hábitat, se asignaron las categorías: Borde del Bosque, Borde del Bosque- Interior del Bosque, Borde del Bosque -No boscoso, Interior del Bosque y No Boscoso, según Stiles (1985), Garrigues y Dean (2007), Stiles y Skutch (2007).
Las especies se clasificaron en cinco categorías, según el nivel de dependencia del bosque (Stiles, 1985; Hidalgo, 2009). En la Clase 1, aves que dependen al menos de un 50% de cobertura boscosa interconectada con grandes áreas de bosque, Clase 1-2, son aquellas que se adaptan a bosques intervenidos. Clase 2, aves que sobreviven en bosques fragmentados y alterados, con menos del 50% de cobertura boscosa. Clase 2-3, aves que necesitan la presencia de árboles asociados a las áreas abiertas y Clase 3, aquellas que no requieren áreas boscosas, pueden sobrevivir en áreas de crecimiento secundario, cercas vivas y áreas de cultivos.
Los individuos se clasificaron según su gremio trófico en, consumidor primario y consumidor secundario (Centeno, 2009; Hidalgo, 2009), a través de la consulta de su historia natural (Garrigues & Dean, 2007; Stiles & Skutch, 2007). Con el fin de clasificar las especies según su condición, se utilizó las categorías determinadas por la lista oficial de especies 2015-2016 de la AOCR.
Cada una de las especies se clasificó de acuerdo a su distribución vertical según Stiles y Skutch (2007). En el estrato bajo, se encuentran los individuos que frecuentan el sotobosque, escarban entre la hojarasca en busca de alimento. En el estrato medio se clasifican todos los individuos que suelen estar en los niveles medios del bosque. Aquellos individuos que acostumbran estar en el dosel del bosque se asignaron en el estrato alto. Finalmente se clasificó en todos los estratos, a todas aquellas especies que se pueden encontrar desde las partes bajas hasta el dosel del bosque.
Para determinar la situación actual de la población se consultó el Reglamento de la Ley de Conservación de la Vida Silvestre (7317), 1992 del MINAE y la Convención sobre el Comercio Internacional de Especies Amenazadas de Flora y Fauna (CITES, 2011), de esta manera se identificaron las especies que pertenecen al Apéndice II, definidas como: especies que no están estrictamente amenazadas de extinción pero en algún momento podrían llegar a estarlo a menos que su comercio se controle estrictamente.
Finalmente se consultó lista roja de especies elaborada por la Unión Internacional para la Conservación de la Naturaleza (IUCN), 2013 con el fin de identificar cuál especie se encontraba como extinta, amenazada y de menor preocupación.
Análisis estadístico: Se utilizó el X 2 simple para evaluar diferencias en las proporciones de especies compartidas entre los diferentes sitios de muestreo (Blake y Loiselle, 2000).
Se realizó la prueba de X2 de contingencia para establecer dependencia entre las variables ecológicas de las especies con el tipo de vegetación. (Fowler y Cohen, 1999). Mediante la ecuación de Clench se determinó el porcentaje de especies registradas en cada sitio, así se logró estimar el registro de especies en los diferentes tipos de vegetación con respecto al esfuerzo de muestreo realizado, con el fin de proponer mejoras metodológicas para estudios futuros (Mao y Chang, 2005; Huitzil, 2007).
RESULTADOS
En el bosque se logró determinar el 84% de las especies seguido de los eucaliptos con un 76% y el pinar con un 48%, (Clench, R2= 0.99).
Se registraron 36 especies pertenecientes a 14 familias en los tipos de vegetación evaluados (Apéndice 1). De éstas, las que presentaron mayor riqueza fueron: Parulidae (n= 7, especies), seguida de la Emberizidae (n= 6), y Trochilidae (n= 4). Existe un alto porcentaje de especies compartidas entre el bosque y los eucaliptos (90%), los eucaliptos y el pinar tienen en común el 63%, sin embargo, entre el bosque y el pinar se comparte un 46,67% (x2= 12,76; p<0.01).
Independientemente del tipo de vegetación, la mayor parte de las especies se caracterizan por encontrarse en el borde del bosque, donde el bosque presentó los valores más altos,17 especies (Cuadro 1). En el pinar los valores difieren solamente por una especie en las categorías borde del bosque y borde del bosque-no boscoso El tipo de hábitat de los individuos resultó ser dependiente del tipo de vegetación (x2= 24,78; p <0.01).
En los tres tipos de vegetación dominan las especies adaptadas a bosques intervenidos; caso contrario sucede con las especies que necesitan menos del 50% de cobertura boscosa, la cual presentó los valores más bajos (Cuadro 2). La clase de dependencia de las aves no está asociada con el tipo de vegetación (x2= 28,14; p>0.10).
El gremio trófico que dominó en todos los tipos de vegetación fueron los consumidores secundarios (Cuadro 3). Por otro lado, el gremio trófico de las especies no depende del tipo de vegetación (x2= 0,95; p>0.10).
Las especies Residentes-Endémicas (Cuadro 4), dominaron en los tres tipos de vegetación (n=15, especies), tanto para bosque como para los eucaliptos. En las plantaciones exóticas se registraron 2 especies bajo la categoría migratoria. Se determinó que la condición de las especies es independiente del tipo de vegetación (x2= 3,04; p>0.10).
En el bosque se registró una mayor riqueza en el estrato medio y alto de la vegetación, al igual que los eucaliptos (n=9); en el caso del pinar se determinó que las especies tienden a encontrarse en el estrato bajo-estrato medio (n=6). La distribución vertical de las especies registradas, está asociada con el tipo de vegetación (x2= 35,46; p<0.01) (Cuadro 5).
Al tomar en cuenta todos los tipos de vegetación y siguiendo la clasificación dada por CITES, se registraron 4 especies que pertenecen al Apéndice II, Colibri cyanotus, Panterpe insignis, Selasphorus flammula, Lampornis calolaemus, todas ellas de la familia Trochilidae, siendo la última, exclusiva del bosque (Apéndice 1).
Ninguna de las especies registradas durante el muestreo se encontró en la lista del Reglamento de la Ley de la Conservación de la Vida Silvestre N° 7317. En cuanto a la lista roja de especies de la IUCN (2013), todas las especies se encontraron bajo la categoría “menor preocupación”. Un taxón es clasificado bajo esta categoría cuando ha sido evaluado bajo los distintos criterios y no clasifica para especies “en peligro crítico, “peligro vulnerable” o “casi amenazado”. Además, se consideran como taxones abundantes y de amplia distribución.
DISCUSIÓN
En este estudio se registró un bajo número de especies, en comparación a estudios realizados en zonas bajas de Costa Rica (Sánchez, 2009). En bosques de elevaciones altas la riqueza declina, debido a la competencia inter e intra específica, cambios en las condiciones ambientales, uso de la tierra, disposición de recursos, entre otros (Young et al., 1998; Blake & Loiselle, 2000; Martínez & Rechberger, 2007).
La composición de la avifauna puede llegar a cambiar en un rango de 10 a 15% por cada 100 m, donde en los sitios de altura las familias más comunes de encontrar son Throchilidae, Parulidae, Thraupidae y Tyrannidae, forrajeando epífitas, vegetación característica de sitios altos (Martínez & Rechberger, 2007). En esta investigación las dos primeras familias mencionadas anteriormente también fueron las más representativas en términos de riqueza, independientemente de los tipos de vegetación.
Según Barlow et al., (2007), las especies que utilizan tanto las plantaciones exóticas y bosques nativos son escasas, en esta investigación existe un alto porcentaje de especies compartidas entre los eucaliptos y el bosque, lo cual concuerda con Paritsis y Aizen (2008), ya que las plantaciones exóticas representan zonas de alimentación, anidamiento y descanso (Parrota, 1995; Collazo & Bonilla, 1988).
La similitud entre las plantaciones exóticas y el bosque nativo puede estar influida por factores como efecto de borde, distancia entre el bosque y la plantación, presencia de un segundo estrato de vegetación, disponibilidad de recursos, tamaño del área, entre otros (Collazo & Bonilla 1988, Marsden et al., 2001, Barlow et al., 2007). Además la distancia que existe entre cada fragmento de vegetación y el bosque natural, favorece el movimiento de aves dentro de estos ecosistemas (Carlson 1986; Parrota, 1995; Collazo & Bonilla, 1988; Paritsis & Aizen, 2008).
Las especies que frecuentan el borde del bosque, utilizan especialmente los árboles de ésta área porque poseen perchas más accesibles y con mejor visibilidad (Parrota, 1995; Geldenhuys, 1997), lo que pudo influir en el mayor registro de aves en esta zona. Las especies generalistas son más abundantes en sitios donde hay un dominio de claros, contrario sucede con las especialistas, que frecuentan el interior del bosque y son más abundantes en bosques continuos (Carlson, 1986; Collazo & Bonilla, 1988; Paritsis & Aizen, 2008; Forsman et al., 2010).
La metodología utilizada durante el muestreo, puede influir en el registro del gremio trófico ya que por medio de observación directa la detección es mayor para especies frugívoras-insectívoras y con la utilización de redes de niebla hay una mayor captura de especies insectívoras de follaje (Blake & Loiselle, 2000; Martínez & Rechberger, 2007).
Barlow et al., (2007) en su investigación difirieron de lo mencionado anteriormente, pues no encontraron diferencias al comparar el tipo de muestreo empleado con el gremio trófico, lo cual apoya los datos encontrados en este estudio.
Las especies insectívoras son comunes en tierras bajas (1850 m.s.n.m.) y raras y ausentes en sitios altos (3170 m.s.n.m.). Además, las especies nectarívoras y frugívoras dominan a medida que aumenta la altitud, debido a una serie de cambios en los recursos disponibles (Martínez & Rechberger, 2007). En este estudio dominaron los consumidores secundarios en la que su dieta se compone principalmente de invertebrados.
La época de fructificación de las especies vegetales limita en cierta parte la presencia de algunos individuos (Loiselle 6 Blake, 1991; Camacho & Orozco, 1998). En el presente estudio esta variable se trató de eliminar mediante la toma de datos tanto en época lluviosa como en época seca. Sumado a lo anterior, se ha demostrado que no existe una correlación entre la cantidad de individuos registrados y la precipitación mensual (Castaño et al., 2008).
El bajo registro de especies migratorias se puede explicar ya que éstas tienden a forrajear en las partes altas y densas del dosel, influyendo en una menor detección, a diferencia de las residentes, comunes en los pisos de la vegetación lo que facilita su avistamiento (Almazán et al., 2009). Por otro lado, en zonas altas es característico encontrar una mayor cantidad de especies endémicas (Martínez & Rechberger, 2007).
La mayor presencia de especies en estratos medios y altos, puede estar influida por la alteración que ha recibido un sitio, aquellos que han sido alterados presentan un estrato intermedio bien desarrollado (Collazo & Bonilla, 1988), con una variedad de microhábitats que atraen a las aves por la diversidad recursos alimenticios, en las áreas no alteradas las especies se distribuyen principalmente en el dosel (Ugalde et al., 2009).
Las plantaciones de pino de la zona se caracterizaban por tener un dosel cerrado, y un sotobosque escaso, a diferencia de los eucaliptos con un dosel de apertura intermedia y un sotobosque dominado por arbustos y plantas similares al bosque (Parrota 1995, Geldenhuys 1997, Cusack & Montagnini 2004). En Costa Rica, estudios indican que los sitios con una apertura del dosel intermedia, presentan una mayor abundancia en la regeneración del sotobosque (Parrota 1995, Geldenhuys 1997, Cusack & Montagnini 2004).
Las especies consideradas como vulnerables se caracterizan por presentar hábitos específicos y poblaciones reducidas, distribuyéndose principalmente en los bosques húmedos (Martínez & Rechberger 2007). No obstante, en el área de estudio, solamente se encontraron 4 especies bajo la categoría II del Apéndice de CITES; las cuales no están estrictamente amenazadas de extinción, pero en algún momento podrían llegar a estarlo.
Las plantaciones pueden funcionar como escalones hacia los bosques naturales aledaños, por lo que pueden ser importantes en la movilidad de las especies (Carlson, 1986; Castaño et al., 2008), como lo son las aves migratorias de áreas abiertas (Carlson 1986, Barlow et al., 2007).
A través de este estudio, se pudo generar información inicial sobre la composición y caracterización ecología de la avifauna en esta zona, necesaria para estudios posteriores donde se recomienda el uso de metodologías convencionales en combinación con nuevas técnicas como lo es la bioacústica, además de incluir la época migratoria, de esta manera aumentará el registro especies (Blake y Loiselle, 2000; Celis et al., 2012).
AGRADECIMIENTOS
A Idea Wild.org, por el apoyo económico, a la UNED, FEES-CONARE por darme la oportunidad de incluirme en su proyecto “Evaluación de factores ecológicos que afectan a la vida silvestre en áreas alteradas y áreas silvestres”, y apoyarme en la logística y transporte en el muestreo piloto, a María Isabel Di Mare, por la motivación de iniciar esta investigación. A Carlos Calvo, Francisco Conejo y Horacio Herrera, por la hospitalidad brindada durante la estadía. A Carmen Hidalgo, Mónica Retamosa, Oscar Ramírez por transmitirme sus conocimientos y sugerencias para el buen desarrollo de la investigación. A Ruperto Quesada por compartir su información referente a la vegetación, a Wilfredo Segura por la ayuda brindada en relación con la geografía del sitio y diseño del mapa. A Javier Rodríguez por las sugerencias para la mejora del manuscrito, a Federico Herrera, Adrián González, Francisco Durán, Jenny Quirós, Jimmy Bravo, ASVO, por la asistencia durante los muestreos.
REFERENCIAS
Almazán, C., Puebla, O., & Almazán, A. (2009). Diversidad de aves en bosques de Pino- encino del centro de Guerrero, México. Acta zoológica Mexicana, 25(19), 123-142.
Asociación Ornitológica de Costa Rica. (2015-2016). Lista oficial de especies. Recuperado de http://listaoficialavesdecostarica.wordpress.com.
Barlow, J., Mestre, L., Gardner, T., 6 Pérez, C. (2007). The value of primary, secondary and plantation forests for Amazonia birds. Biological Conservation, 136, 212-231. doi: 10.1016/j.biocon.2006.11.021
Bermúdez, F., & Conejo, R. (2008). Sistema Nacional de Áreas de Conservación: Área de Conservación Cordillera Volcánica Central. Plan Maestro. Parque Nacional Volcán Irazú. 2-105 .
Blake, J., & Loiselle, B.(2000). Diversity of Birds Along and Elevational Gradient in the Cordillera Central, Costa Rica. The Auk, 117,663-686. doi: 10.2307/4089592
Blake, J. (1992). The temporal variation in point counts of birds in a lowland wet forest in Costa Rica. The Condor ,9, 265-267. doi: 10.2307/1368816
Botero, E. (2005). Métodos para estudiar las aves. Biocarta. Programa Biología de la Conservación. Cenicafé,8,1-4.
Camacho, M., & Orozco, L. (1998). Patrones fenológicos de doce especies arbóreas del bosque montano de la Cordillera de Talamanca, Costa Rica. Revista de. Biología. Tropical,46(3), 533-542.
Carlson, A. (1986). A comparison of birds inhabiting pine plantation and indigenous forest patches in a tropical mountain area. Biological Conservation, 35(3),195–204. doi: 10.1016/0006-3207(86)90063-7
Castaño, G., y Patiño, J. C. (2000). Cambios en la composición de la avifauna en Santa en Helena durante el siglo XX. Crónica forestal y del medio ambiente,15(1), 1-2.
Castaño, J., Morales, J. A., & Bedoya, M. L. (2008). Aportes de una plantación forestal mixta a la conservación de la Avifauna en el Cañon del Río Cauca, Colombia.Revista Faultad Nacional de Agronomía Medellín, 61(1), 4358-4365.
Celis, A., Deppe, J., & Ward, M. (2012). Effectiveness and utility of acoustic recordings for surveying tropical birds. Journal field of ornithology. doi: 10.1111/j.1557-9263.2012.00366.x
Centeno, M. S. (2009). Caracterización ecológica la comunidad avifaunística de la Escuela de Agricultura de la Región del Trópico Húmedo-La Flor, Liberia, Guanacaste (Tesis de pregrado). Universidad Nacional,Heredia, Costa Rica.
Cerezo, A., Robbins, C., & Dowell B. (2009). Uso de hábitats modificados por aves dependientes de bosque tropical en la región caribeña de Guatemala. Revista de Biología Tropical, 57(1-2), 401- 419.
CITES. (2011). Apéndices I, II y III. Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres Recuperado de: http://www.cites.org/esp/app/index.shtml.
Collazo, A., & Bonilla, G. (1988). Comparación de la riqueza de aves entre plantaciones de pino hondureño (Pinus caribaea) y áreas de bosque nativo en el Bosque Estatal de Carite, Cayey, Puerto Rico. Caribbean journal of Science, 24(1-2), 1- 10.
Colwell, R. ( 2009). EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples,Software and (User´s Guide). Recuperado de http://viceroy.eeb.uconn.edu/estimates.
Cusack, D., & Montagnini, F. (2004). The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica. Forest Ecology and Management 188, (1-3), 1-15. doi: 10.1016/S0378-1127(03)00302-5
Fonseca, P. (2009). Prusia cambia pinos, eucaliptos y cipreses por especies locales. La Nación. Recuperado de http://www.nacion.com/vivir/Prusia-eucaliptos-cipreses-especies-locales_0_1043095758.html.
Forsman, J., Reunanen, P., Jokimäki, J., & Iviönkkönen, M. (2010). The effects of small-scale disturbance on forest birds: a meta-analysis. Canadian Journal of Forest Research, 40(9), 1833-1842. doi: 10.1139/X10-126
Fowler, J., & Cohen, L. (1999). Estadística Básica en Ornitología .Madrid, España: Seo/ BirdLife.
Garrigues, R., & Dean, Y. (2007). The Birds of Costa Rica. Cornell University: Ithaca, New York:
Geldenhuys, J. (1997).Pine and eucalypt plantations in Northern Province, South Africa. Forest Ecology and Management, 99(1-2), 101–115. doi: 10.1016/S0378-1127(97)00197-7
Guariguata, M., & Ostertag, R.( 2001). Neotropical secondary forest succession: changes in structural and functional characteristics. Forest Ecology and Management, 148, 185-206.
Hidalgo, C. (2009). Principales variables ecológicas para el manejo y la caracterización de la avifauna costarricense. Poligrafiado. Universidad Nacional, Heredia, Costa Rica.
Hobbs, R., Catling, P. C., Wombey, J. C., Clayton, M., Atkins, L., & Reid, A. (2003). Faunal use of bluegum (Eucalyptus globulus) plantations in southwestern Australia. Agroforestry Systems, 58, 195–212. doi: 10.1023/A:1026073906512
Huitzil, J. C. (2007). Herpetofauna de dos localidades de la región Norte de Zimapán, Hidalgo. (Tesis pregrado). Universidad Autónoma del estado de Hidalgo, Hidalgo, México.
International Union for Coservation of Nature and Natural Resources (IUCN). (2013).The IUCN the Red List 2013. Recuperado de http://www.iucnredlist.org/amazing-species.
Loiselle, A., & Blake, J. (1991). Temporal variation in birds and fruits along an elevational gradient in Costa Rica, Ecology 72 (1), 180-193. doi: 10.2307/1938913
Marsden, S., Whiffin, M., & Galetti, M. (2001). Bird diversity and abundance in forest fragments and Eucalyptus plantations around an Atlantic forest reserve, Brazil. Biodiversity and Conservation,10, (5), 737–751. doi: 10.1023/A:1016669118956
Martínez, O., & Rechberger, J. (2007). Características de la avifauna en un gradiente altitudinal de un bosque nublado andino en La Paz, Bolivia. Revista Peruana de Biología, 14 (2), 225-236.
Martínez, S. (2008). Conectividad funcional para las aves terrestre en un paisaje fragmentado en Mantiguás, Nicaragua (Tesis de maestría) CATIE, Turrialba, Costa Rica.
Mao, C., & Chang, J. (2005). Interpolando, extrapolando y comparando las curvas de especies basadas en su incidencia. Sobre Diversidad Biológica: El significado de las Diversidades Alfa, Beta y Gamma. Monografías Tercer Milenio, 4,73-84.
MINAE. (1992). Reglamento de la Ley de Conservación de la Vida Silvestre. Número 7317. Dirección Regional de Vida Silvestre, Ministerio de Ambiente y Energía (MINAE). La Uruca, San José, Costa Rica: La Gaceta.
Murata, K. J., Dondoli, C., &. Sáenz, R. (1966). The 1963–65 eruption of Irazú Volcano, Costa Rica (the period of March 1963 to October 1964). Bulletin Volcanologique,29 (1), 763-793. doi: 10.1007/BF02597194
Niehaus, C., Valerio, I., Blanco, K., & Chinchilla, M. (2012). Infecciones parasitarias del coyote, Canis latrans (Carnivora: Canidae). Revista de Biología. Tropical, 60 (2), 799-808. doi: 10.15517/rbt.v60i2.4000
Paritsis, J., & Aizen, M. (2008). Effects of exotic conifer plantations on the biodiversity of understory plants, epigeal beetles and birds in Nothofagus dombeyi forests. Forest Ecology and Management, 255 (5-6),1575–1583. doi: 10.1016/j.foreco.2007.11.015
Parrota, J. (1995). Influence of overstory composition on understory colonization by native species in plantations on a degraded tropical. Journal of Vegetation Science, 6 (5), 627-636. doi: 10.2307/3236433
Quesada, R. (2007). IX Congreso Nacional de Ciencias, exploraciones fuera y dentro del aula: Los Bosques de Costa Rica. Cartago, Costa Rica: Instituto Tecnológico de Costa Rica.
Ralph, J., Geupel, G., Pyle, P., Martin, T., DeSante, D., & Milá, B. (1996). Manual de Métodos de campo para el monitoreo de aves terrestres. California, Estados Unidos: Pacific Southwest Research Station. doi: 10.2737/PSW-GTR-159
Ramírez, J. (2010). Diversidad de aves de hábitats naturales y modificados en un paisaje de la Depresión Central de Chiápas, México. Revista de Biología Tropical, 58 (1),511-528.
Sánchez, A. (2009). Ensamble y ocurrencia de aves en bosques sometidos a manejo forestal, Región Huetar Note, Costa Rica (Tesis de Posgrado). Universidad Nacional. Heredia, Costa Rica.
Sánchez, O., Zamorano, P., Peters, E., & Moya, H. (2011). Temas sobre la conservación de vertebrados silvestres en México. Calzada de San Lorenzo, México: Secretaría de Medio Ambiente y Recursos Naturales.
Santos, T., & Tellería, J. L. (2006). Pérdida y fragmentación del hábitat: efecto sobre la conservación de las especies. Ecosistemas, 15 (2), 3-12.
Stiles, F. (1985). Conservation of forest birds in Costa Rica: Problems and Perspectives. Council Bird: Cambridge, Inglaterra:
Stiles, F., & Skutch, A. (2007). Guía de aves de Costa Rica. INbio: Santo Domingo, Heredia.
Ugalde, S., Valdez, J. I., Hernandéz, G., Ramírez, G., & Alcántar, J. l. (2009). Distribución vertical de aves en un bosque templado con diferentes niveles de perturbación Madera y Bosques, 15 (1), 5-26. doi: 10.21829/myb.2009.1511194
Young, B., Derosier, D., & Powell, G. (1998). Diversity and Conservation Understory Birds in the Tilarán Mountains, Costa Rica. The Auk, 115(4), 998-1016. doi: 10.2307/4089518
RESUMEN: El sector Prusia del Parque Nacional Volcán Irazú, Costa Rica, tiene plantaciones de especies nativas y exóticas que incluyen pino, ciprés y eucalipto. Estimé la riqueza de aves con puntos de conteo (radio fijo), en las plantaciones de pino, eucalipto y de bosque, se encontraron 36 especies distribuidas en 14 familias. El eucalipto y el bosque comparten el 90% de las especies. El tipo de hábitat y la distribución vertical de las especies fueron las únicas dos variables que mostraron estar asociadas al tipo de vegetación. La baja riqueza de especies encontradas en este estudio puede explicarse a la zona altitudinal y la homogeneidad de los fragmentos de vegetación exótica que podrían estar causando una disminución de recursos alimenticios y sitios de refugio, mientras que la falta de asociaciones ecologías de las especies con el tipo de vegetación, podrían ser el resultado de la distancia de la reserva natural, presencia de un estrato medio de vegetación, o algunos otros factores.
Palabras clave: Riqueza, aves, Costa Rica, bosque, plantaciones exóticas.

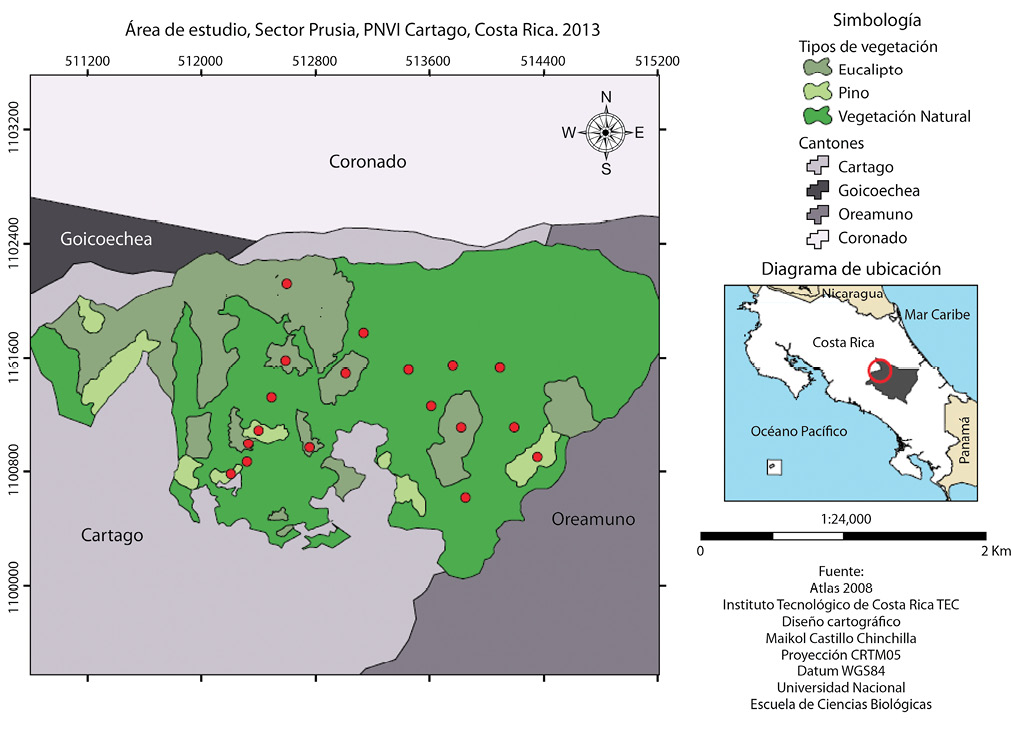
Fig. 1. Área de estudio en el Sector de Prusia, del Parque Nacional Volcán Irazú, Cartago, Costa Rica. Los puntos rojos representan algunos sitios de muestreos en los distintos tipos de vegetación estudiados (Fuente: Castillo, 2014, diseño del mapa. Segura, 2009, capas de uso del suelo del sector Prusia).
CUADRO 1
Riqueza de especies de aves según el tipo de hábitat encontrado en los tres tipos de vegetación
|
Tipo de hábitat |
|||||
|
Vegetación |
Borde del Bosque |
Borde del Bosque- Interior del Bosque |
Borde del Bosque- No boscoso |
Interior del Bosque |
No Boscoso |
|
Bosque |
17 |
0 |
12 |
0 |
1 |
|
Pinar |
9 |
0 |
8 |
0 |
0 |
|
Eucaliptos |
16 |
1 |
9 |
1 |
0 |
CUADRO 2
Clase de dependencia del bosque de las especies registradas en los tres tipos de vegetación
|
Tipo de vegetación |
Clase 1: Dependen del 50% cobertura |
Clase 1-2: Adaptadas a vivir en bosques intervenidos |
Clase 2 Necesitan menos del 50% de cobertura boscosa |
Clase 2-3: Requieren presencia de árboles de áreas abiertas |
Clase 3: No requieren áreas boscosas |
|
Bosque |
1 |
16 |
1 |
5 |
7 |
|
Pinar |
1 |
8 |
0 |
4 |
4 |
|
Eucaliptos |
3 |
13 |
1 |
6 |
4 |
CUADRO 3
Clasificación de las especies según su gremio trófico en los diferentes tipos de vegetación
|
|
Gremio trófico |
|
|
Tipo de vegetación |
Consumidor Primario |
Consumidor Secundario |
|
Bosque |
9 |
21 |
|
Pinar |
4 |
13 |
|
Eucaliptos |
7 |
20 |
CUADRO 4
Condición de las especies registradas en los diferentes tipos de vegetación
|
Condición de la especie |
|||
|
Tipo de vegetación |
Migratorio |
Residente |
Residente-Endémica |
|
Bosque |
4 |
11 |
15 |
|
Pinar |
2 |
7 |
8 |
|
Eucaliptos |
2 |
10 |
15 |
CUADRO 5
Distribución vertical de las especies asociadas a cada tipo de vegetación
|
Distribución vertical de las especies |
|||||||
|
Tipo de vegetación |
Estrato Alto |
Estrato Alto - Estrato Bajo |
Estrato Bajo |
Estrato Bajo - Estrato Medio |
Estrato Medio |
Estrato Medio - Estrato Alto |
Todos los estratos |
|
Bosque |
1 |
1 |
7 |
4 |
1 |
9 |
7 |
|
Pinar |
0 |
1 |
3 |
6 |
0 |
4 |
3 |
|
Eucaliptos |
1 |
1 |
7 |
2 |
1 |
9 |
6 |
APÉNDICE 1
Riqueza y caracterización de las especies de aves encontradas en el Sector de Prusia del Parque Nacional Volcán Irazú.
|
Especies encontradas en cada sitio |
Situación actual de la población |
|||||||||||
|
Familia |
Especies |
Nombre común en inglés |
BN |
P |
E |
Tipo de Hábitat |
Clase de dependencia del bosque |
Gremio trófico |
Condición de la especie |
Distribución vertical |
Apendice CITES |
UICN Lista Roja: Criterios y Categorías |
|
Odonthoporidae |
Odontophorus guttatus |
Spotted Wood-Quail |
x |
IB |
1 |
I |
R |
EB |
LC |
|||
|
Columbidae |
Patagioenas fasciata |
Band-tailed Pigeon |
x |
x |
BB |
2 |
I |
R |
EA |
LC |
||
|
Trochilidae |
Colibri cyanotus |
Lesser Violetear |
x |
x |
x |
BB-NB |
2-3 |
I |
R |
EM-EA |
II |
LC |
|
Trochilidae |
Panterpe insignis |
Fiery-throated Hummingbird |
x |
x |
x |
BB |
2 |
I |
R-END |
EM-EA |
II |
LC |
|
Trochilidae |
Lampornis calolaemus |
Purple - throated Mountain - gem |
x |
BB |
2 |
I |
R-END |
EM-EA |
II |
LC |
||
|
Trochilidae |
Selasphorus flammula |
Volcano Hummingbird |
x |
x |
x |
BB-NB |
3 |
I |
R-END |
EM-EA |
II |
LC |
|
Picidae |
Melanerpes formicivorus |
Acorn Woodpecker |
x |
BB-NB |
2 |
I |
R |
TE |
LC |
|||
|
Picidae |
Picoides villosus |
Hairy Woodpecker |
x |
x |
x |
BB-NB |
2 |
II |
R |
TE |
LC |
|
|
Furnariidae |
Lepidocolaptes affinis |
Spot-crowned Woodcreeper |
x |
x |
BB |
2 |
II |
R |
EM-EA |
LC |
||
|
Furnariidae |
Margarornis rubiginosus |
Ruddy Treerunner |
x |
x |
BB |
2 |
II |
R-END |
TE |
LC |
||
|
Furnariidae |
Cranioleuca erythrops |
Red - faced Spinetail |
x |
x |
x |
BB |
2 |
II |
R |
EB-EM |
LC |
|
|
Tyrannidae |
Elaenia frantzii |
Mountain Elaenia |
x |
x |
x |
BB-NB |
2-3 |
II |
R |
EA-EB |
LC |
|
|
Tyrannidae |
Empidonax atriceps |
Black - capped Flycatcher |
x |
BB |
2-3 |
II |
R-END |
EM-EA |
LC |
|||
|
Vireonidae |
Vireo carmioli |
Yellow-winged Vireo |
x |
x |
BB |
2 |
II |
R-END |
TE |
LC |
||
|
Troglodytidae |
Troglodytes aedon |
House wren |
x |
NB |
3 |
II |
R |
EB |
LC |
|||
|
Troglodytidae |
Henicorhina leucophrys |
Gray-breasted Wood-Wren |
x |
x |
BB |
1 |
II |
R |
EB |
LC |
||
|
Turdidae |
Catharus gracilirostris |
Black-billed Nightingale-Thrush |
x |
x |
BB |
2 |
II |
R-END |
TE |
LC |
||
|
Turdidae |
Catharus frantzii |
Ruddy - capped Nightingale - Trush |
x |
x |
BB |
2 |
II |
R |
EB |
LC |
||
|
Turdidae |
Turdus nigrescens |
Sooty Thrush |
x |
x |
x |
BB |
3 |
II |
R-END |
EB |
LC |
|
|
Ptiliogonidae |
Phainoptila melanoxantha |
Black-and-Yellow Silky-Flycatcher |
x |
BB |
2 |
I |
R-END |
EM |
LC |
|||
|
Ptiliogonidae |
Ptiliogonys caudatus |
Long-tailed Silky-Flycatcher |
x |
x |
x |
BB |
2-3 |
II |
R-END |
EM-EA |
LC |
|
|
Parulidae |
Vermivora cyanoptera |
Blue- winged Warbler |
x |
BB-NB |
3 |
II |
M |
EM |
LC |
|||
|
Parulidae |
Oreothlypis gutturalis |
Flame-throated Warbler |
x |
x |
BB |
2 |
II |
R-END |
EM-EA |
LC |
||
|
Parulidae |
Setophaga virens |
Black-throated Green Warbler |
x |
x |
x |
BB-NB |
2 |
II |
M |
EM-EA |
LC |
|
|
Parulidae |
Basileuterus melanogenys |
Black-cheeked Warbler |
x |
x |
BB |
1-2 |
II |
R-END |
EB |
LC |
||
|
Parulidae |
Cardellina pusilla |
Wilson’s Warbler |
x |
x |
x |
BB-NB |
2-3 |
II |
M |
TE |
LC |
|
|
Parulidae |
Myioborus miniatus |
Slate - throated Redstart |
x |
BB |
2 |
II |
R |
EB-EM |
LC |
|||
|
Parulidae |
Myioborus torquatus |
Collared Redstart |
x |
x |
BB |
2 |
II |
R-END |
EB-EM |
LC |
||
|
Emberizidae |
Diglossa plumbea |
Slaty Flower-piercer |
x |
x |
BB-NB |
2-3 |
I |
R-END |
EM-EA |
LC |
||
|
Emberizidae |
Pselliophorus tibialis |
Yellow-thighed Finch |
x |
x |
x |
BB-NB |
2 |
II |
R-END |
TE |
LC |
|
|
Emberizidae |
Pezopetes capitalis |
Large-footed Finch |
x |
x |
x |
BB |
2 |
II |
R-END |
EB |
LC |
|
|
Emberizidae |
Arremon brunneinucha |
Chestnut-capped Brush - Finch |
x |
BB-IB |
1 |
II |
R |
EB |
LC |
|||
|
Emberizidae |
Zonotrichia capensis |
Rufous-collared Sparrow |
x |
x |
BB-NB |
3 |
I |
R |
EB-EM |
LC |
||
|
Emberizidae |
Chlorospingus pileatus |
Sooty-capped Bush-tanager |
x |
x |
x |
BB |
3 |
II |
R-END |
EB-EM |
LC |
|
|
Cardinalidae |
Pheucticus ludovicianus |
Rose - breasted Grosebeak |
x |
BB-NB |
3 |
I |
M |
EB-EA |
LC |
|||
|
Icteridae |
Amblycercus holosericeus |
Yellow-billed Cacique |
x |
BB-NB |
3 |
II |
R |
EB |
LC |
|||
ABREVIATURAS
Especies encontradas en cada sitio
B: bosque
P: pinar
E: eucaliptos
Tipo de hábitat
BB: Borde del Bosque
NB: No Boscoso
IB: Interior del Bosque
Clase de dependencia del bosque
Clase 1: Se encuetran todas aquellas especies que dependen al menos de un 50% de la cobertura boscosa interconectada con grandes áreas de bosque.
Clase 1-2: Son las especies se adaptan a bosques intervenidos.
Clase 2: Son las especies que están adaptadas a sobrevivir en bosques considerados como fragmentados y alterados, los cuales poseen menos del 50% de cobertura boscosa.
Clase 2-3: Especies que necesitan por lo menos la presencia de árboles asociados a las áreas abiertas.
Clase 3: En esta categoria están las especies que no requieren áreas boscosas, ya que pueden sobrevivir en áreas de crecimiento secundario, cercas vivas y áreas de cultivos.
Gremio trófico
I: consumidor primario: incluye organismos que se alimentan de: Frutos-insectos (incluye artrópodos pequeños, frutos-semillas (ariladas) - néctar-parte vegetativo e invertebrado, frutos-semillas (ariladas) - partes vegetativas, néctar-insectos, arañas y Semillas de gramíneas-partes vegetativas-insectos-frutos.
II: consumidor secundario: se clasifican es estas categorías las aves que su dienta se basa en el consumo de: insectos, insectos grandes-frutos-semillas ariladas, insectos-frutos-semillas (ariladas)-néctar-partes vegetativas e insectos-otros invertebrados pequeños (arañas).
Condición de la especie:
R: residente, especie que se podría encontrar durante cualquier época del año en Costa Rica y además su reproducción ha sido comprobada.
M: migratorio, especie que no se ha comprobado su reproducción en Costa Rica, sin embargo pasa parte del año en el territorio, además existen individuos no reproductivos que pueden encontrarse durante todo el año en el país.
END: especies endémicas, aquellas especies que se encuentran únicamente en Costa Rica y la Región.
Distribución vertical de las especies:
EB: estrato bajo, individuos que se encuentran en el sotobosque, escarban entre la hojarasca en busca de alimento.
EM: estrato medio, individuos que frecuentan los niveles medios del bosque.
EA: estrato alto, individuos que frecuentan el dosel del bosque
TE: todos los estratos, especies que se pueden encontrar desde las partes bajas hasta el dosel del bosque.
Situación actual de la población
II: Apéndice II de CITES, se consideran en esta categoría a las especies que no están estrictamente amenazadas de extinción pero en algún momento podrían llegar a estarlo a menos que se su comercio se controle estrictamente.
LC: según clasificación de la IUCN se consideran bajo esta categoría a las especies de menor preocupación y no clasifican en las categorías de peligro crítico, en peligro vulnerable o casi amenazado. Además se consideran como taxones abundantes y de amplia distribución.