Biocenosis • ISSN: 1659-4991/ Vol. 36/ Núm. 2/ Julio-Diciembre, 2025
Interacciones colibrí–planta en jardines de alta montaña: estudio de caso en Paraíso Quetzal Lodge y finca Doña Blanca (Los Santos, Costa Rica, 2024)
Hummingbird–plant interactions in high-mountain gardens: a case study at Paraíso Quetzal Lodge and finca Doña Blanca (Los Santos, Costa Rica, 2024)
Rebeca Navarro Araya1
Paola Brenes Rojas2
Catalina Vargas Meneses3
DOI: 10.22458/rb.v36i2.6273
Recibido – Received: 08 / 06 / 2025 / Corregido – Revised: 18 / 10 / 2025 / Aceptado – Accepted: 20 /10 / 2025
RESUMEN
Se evaluaron las interacciones colibrí–planta en dos jardines de alta montaña en Los Santos, Costa Rica (Paraíso Quetzal Lodge y finca Doña Blanca; 2 650–2 700 m), durante abril–junio de 2024, mediante observación estandarizada en parcelas (ventanas de 10 min con repeticiones mañana/tarde). Se registraron siete especies de colibríes y se caracterizó la composición florística por sitio. La visitación se concentró en un subconjunto de plantas con morfología compatible y oferta sostenida de néctar, principalmente Streptosolen jamesonii (arbusto mermelada), Stachytarpheta mutabilis (rabo de gato), Fuchsia spp. (fucsia) y Myrcianthes storkii (guayabillo), mientras que varias plantas ornamentales mostraron interacción marginal o nula. La proximidad de perchas (arbustos/árboles) y la estratificación vertical del jardín se asociaron con mayor permanencia y retorno de las aves, y explican diferencias entre un sitio consolidado y otro en consolidación. A partir de estos patrones, se proponen lineamientos de diseño: composición centrada en 2–3 especies núcleo de floración sostenida, inclusión de una planta “nodo”, incorporación de elementos de percha cercanos, distancias cortas entre “islas de néctar” y perchas, bloques densos y manejo fenológico escalonado (estrategia agrícola que consiste en programar y distribuir las etapas de floración, fructificación o cosecha de una planta en el tiempo). Los resultados ofrecen una base operativa para fortalecer la conservación de colibríes y el aviturismo local en ambientes de alta montaña.
Palabras clave: conservación de colibríes; perchas (microhábitats); jardines polinizadores; diseño ecológico; aviturismo.
ABSTRACT
Hummingbird–plant interactions were assessed in two high‐mountain gardens in Los Santos, Costa Rica (Paraíso Quetzal Lodge and finca Doña Blanca; 2,650–2,700 m) in April–June 2024 using standardized plot observations (10-min windows with morning/afternoon repetitions). Seven hummingbird species were recorded, and garden floristic composition was characterized. Visitation concentrated on a reduced set of nectar-rich, morphologically compatible plants—primarily Streptosolen jamesonii, Stachytarpheta mutabilis, Fuchsia spp., and Myrcianthes storki, while several ornamentals showed marginal or no interaction. Proximity of perches (shrubs/trees) and vertical stratification were associated with longer stays and returns of the birds, which explains differences between a consolidated and an emerging garden. These patterns allow the following design guidelines: 2–3 core species with sustained flowering, one “node plant,” nearby perches, short distances between nectar “islands” and perches, dense blocks, and staggered phenological management (an agricultural strategy that involves scheduling and spreading the stages of flowering, fruiting, or harvesting of a plant over time). The findings provide an operational basis to strengthen hummingbirds conservation and local birdwatching in high-mountain settings.
Key words: hummingbird conservation; perches (microhabitats); pollinator gardens; ecological design; birdwatching tourism.
Introducción
La interacción entre los colibríes y las plantas polinizadoras en la zona alta de Los Santos, Costa Rica, representa un ejemplo de coevolución en el que ambos grupos de especies obtienen beneficios para su supervivencia. En este contexto, los colibríes no solo dependen del néctar producido por las flores para su nutrición, sino que también realizan la polinización de las plantas, garantizando su reproducción. Las características del pico de los colibríes (forma y longitud) y la estructura de las flores, especialmente las corolas tubulares, son señales de esta adaptación mutua (Burbano-Álvarez, 2013; Amaya et al., 2001).
Esta relación, sin embargo, se ve modulada por factores antropogénicos y ecológicos, como uso del suelo, disponibilidad de recursos y competencia interespecífica, que condicionan la presencia y la actividad de los colibríes en ecosistemas de montaña (Rodríguez & Stiles, 2005). Organismos internacionales como la Plataforma Intergubernamental Científico Normativa sobre Diversidad Biológica y Servicios de los Ecosistemas han advertido sobre la pérdida de polinizadores y la urgencia de implementar estrategias de conservación; por ello, los jardines para polinizadores emergen como herramientas visibles y aplicables para sostener interacciones clave, promover educación y fortalecer la sostenibilidad territorial (IPBES, 2016; Faigón, 2021). En Los Santos, estos jardines además catalizan turismo científico y conservación de especies, siempre que la selección de plantas se realice con criterios técnico-científicos y se documenten las interacciones para mejorar la eficacia del manejo.
La pertinencia territorial es evidente: la región alta de Los Santos con oferta ecoturística, corredores biológicos y paisajes boscosos, concentra un creciente interés por el aviturismo y el establecimiento de jardines de observación de aves. No obstante, carece de estudios específicos que orienten cuáles especies y de qué manera disponerlas espacialmente resulta más adecuado para promover la visitación deseada. Esta brecha operativa limita el crecimiento y la efectividad de las iniciativas locales.
En respuesta, la reciente Guía metodológica para jardines de observación de colibríes para Los Santos (Brenes, et al., 2024) enfatiza en los siguientes puntos: (i) el éxito de estos jardines depende de floración continua y de considerar factores climáticos al elegir especies; (ii) una metodología en 12 pasos que prioriza plantas nativas, creación de áreas de descanso/perchas y jardinería orgánica; así como (iii) la conveniencia de iniciar con especies accesibles de comprobada atracción para colibríes.
Mediante esta investigación se alcanzó el siguiente paso: evaluar las interacciones colibrí–planta en jardines representativos de alta montaña en Los Santos e interpretar los patrones en reglas de diseño verificables (composición, estratificación y manejo) aplicables a los contextos locales. Específicamente, se caracterizó la composición florística y los elementos de microhábitat (perchas/refugio); además, se cuantificó la visitación por especie de planta y por sitio, y se propusieron lineamientos prácticos para jardines polinizadores orientados a la conservación y al aviturismo local, en coherencia con el enfoque técnico-científico recomendado por la guía.
Bajo este marco, se plantea que: (i) las plantas con corola tubular y floración sostenida exhibirán mayores tasas de visita de colibríes, en coherencia con la coevolución morfológica (Burbano-Álvarez, 2013; Amaya et al., 2001); (ii) la proximidad a perchas/arbustos incrementará la actividad de forrajeo al reducir costos energéticos asociados a defensa y descanso; y (iii) los jardines con estratificación vertical (herbáceas–arbustos–árboles) y distancias funcionalmente cortas entre núcleos nectaríferos y perchas concentrarán la interacción en escenarios de competencia de montaña (Rodríguez & Stiles, 2005). En conjunto, el estudio aporta reglas de diseño contextualizadas para Los Santos, las cuales actores locales (propietarios, centros educativos, emprendimientos) pueden adoptar (alineadas con criterios técnico-científicos, documentación de interacciones y condicionantes edafoclimáticos) con el fin de fortalecer conservación, educación ambiental y rutas de avistamiento en la región.
Materiales y métodos
La investigación se llevó a cabo en el cantón de Dota, provincia de San José, Costa Rica. Zona clasificada como bosque muy húmedo montano bajo, clima fresco y templado, con una vegetación densa y rica en biodiversidad. Se seleccionaron dos sitios específicos: finca Doña Blanca, ubicada en Jardín de Dota a una altitud de 2700 m.s.n.m. (9°42´39” N y 83°57´59” O) -figura 1- y Paraíso Quetzal Lodge, en San Gerardo de Dota a 2650 m s.n.m. (9°38´38.76”» N y 83°51´1.08” O) -figura 2-.
Figura 1
Se muestra la ubicación de la finca Doña Blanca y de las parcelas delimitadas dentro de la zona
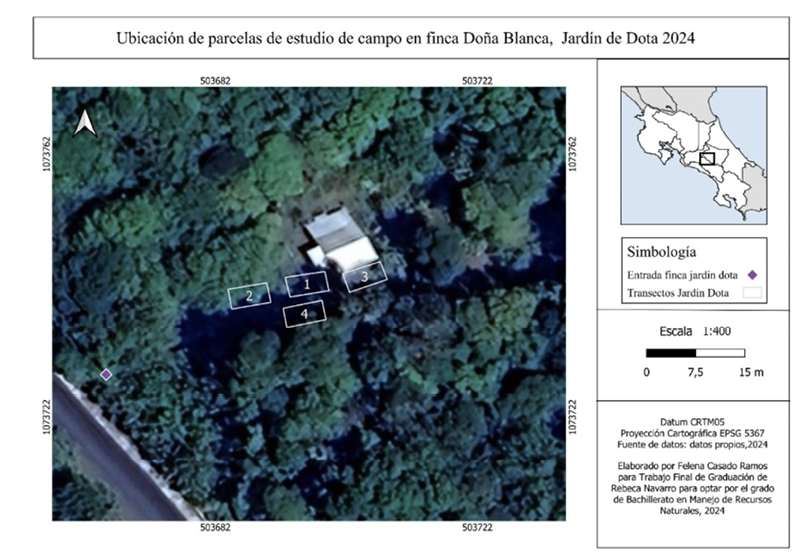
Nota. La figura muestra la ubicación geográfica junto con las parcelas establecidas para la toma de datos
en la finca Doña Blanca, a través de QGIS como sistema de información geográfica.
Figura 2
Ubicación Paraíso Quetzal Lodge y parcelas delimitadas dentro de la zona

Nota. La figura muestra la ubicación geográfica junto con las parcelas establecidas para la toma de datos
en Paraíso Quetzal Logde, a través de QGIS como sistema de información geográfica.
Los sitios se seleccionaron por su potencial: Paraíso Quetzal es un referente regional en aviturismo, mientras que Doña Blanca es un espacio en consolidación para conservación y educación; ambos se ubican en paisajes de robledales de altura con oportunidades para el ecoturismo local. Durante el muestreo, Doña Blanca (abril) presentó mañanas soleadas con transición a nubosidad parcial y lloviznas, en tanto que Paraíso Quetzal (mayo–junio) registró condiciones mayormente soleadas. Las figuras 1 y 2 muestran la localización y el esquema de muestreo cartografiado en QGIS (CRTM05/EPSG:5367).
En cada sitio, se demarcaron cuatro parcelas rectangulares de 6 × 3 m con estacas de madera y cinta métrica, dispuestas como réplicas espaciales sin solapamiento. En cada parcela se levantó el inventario florístico y se registraron las especies de colibríes observadas; la identificación se apoyó en fotografías de campo, consulta a especialistas y verificación en fuentes en línea cuando fue pertinente.
El trabajo de campo comprendió tres visitas por sitio durante 2024. En la finca Doña Blanca se muestreó los días 6, 20 y 28 de abril; mientras en Paraíso Quetzal, los días 11 y 25 de mayo y el 8 de junio. En cada fecha se recorrieron todas las parcelas en dos periodos (mañana y tarde; 08:30 a.m.–12:30 p.m.) y la actividad se registró en ventanas fijas de 10 minutos con tres repeticiones por parcela y por periodo, anotando la visitación por especie de planta y los comportamientos asociados (percha, alimentación, territorialidad). Con este protocolo, el esfuerzo mínimo por visita equivalió a 60 min por parcela (dos periodos × tres repeticiones × 10 min), 240 minutos por sitio y visita, y 720 minutos acumulados por sitio en toda la campaña.
Los datos de visitación se organizaron por especie de planta, parcela y sitio, y se analizaron mediante estadística descriptiva para identificar patrones comparativos de interacción colibrí-planta. La cartografía de apoyo se generó en QGIS (escala 1:400; CRTM05/EPSG:5367) a partir de datos propios obtenidos en 2024. Los lineamientos de diseño que se discuten posteriormente se contrastan con la Guía metodológica para el desarrollo de jardines de observación de colibríes para la zona de Los Santos con el fin de adaptar la selección de especies, la disposición espacial (incluida la estratificación y la ubicación de perchas) y el manejo a las condiciones locales (Brenes, Vargas y Muñoz, 2024).
El estudio se basó en observación no extractiva y sin manipulación de fauna o flora, con el permiso de los propietarios de ambos sitios.
Composición florística de las parcelas en jardines de visitación
Se registró un total de siete especies de colibríes para ambos sitios de estudio: Selasphorus flammula, Lampornis calolaemus, Lampornis castaneoventris, Selasphorus scintilla, Panterpe insignis, Eugenes spectabilis y Colibri cyanotus, todas pertenecientes a la familia Trochilidae y distribuidas en ambos sitios de estudio, a excepción de Eugenes spectabilis; observada solamente en Paraíso Quetzal.
Las cuatro parcelas evaluadas para la finca Doña Blanca evidenciaron diferencias en la riqueza y diversidad florística. Se observaron cinco especies en la parcela 1, tres en la parcela 2, dos en la parcela 3 y una en la parcela 4, tal como se muestra en la tabla 1. Las especies de plantas más comunes: Streptosolen jamesonii (arbusto mermelada), con individuos en todas las parcelas, y Stachytarpheta mutabilis (rabo de gato), registrada en la parcela 1. La diversidad de especies presentó una disminución, alcanzando su punto más bajo en la parcela 4 donde no se registró ningún individuo.
Tabla 1
Riqueza, abundancia y diversidad de plantas por parcela en finca Doña Blanca (Jardín de Dota, Dota, San José)
|
Especie de planta |
Parcela 1 |
Parcela 2 |
Parcela 3 |
Parcela 4 |
|
Psidium cattleianum (guayabita del Perú) |
1 |
0 |
0 |
0 |
|
Agapanthus africanus (agapanto africano) |
1 |
0 |
0 |
0 |
|
Stachytarpheta mutabilis (rabo de gato) |
2 |
0 |
0 |
0 |
|
Streptosolen jamesonii (arbusto mermelada) |
3 |
2 |
2 |
3 |
|
Eriobotrya japonica (níspero) |
1 |
1 |
0 |
0 |
|
Strelitzia reginae (ave del paraíso) |
0 |
1 |
0 |
0 |
|
Tibouchina urvilleana (arbusto de la gloria) |
0 |
0 |
1 |
0 |
|
Total de individuos |
8 |
4 |
3 |
3 |
|
Riqueza de especies |
5 |
3 |
2 |
1 |
|
Índice diversidad |
1.494 |
1.040 |
0.637 |
0.000 |
Por otro lado, en Paraíso Quetzal, aunque todas las parcelas presentaron variaciones en la totalidad de individuos, mostraron una riqueza similar por parcela, como se observa en la tabla 2. Se registró la presencia de Fuchsia spp. (fucsias), en tres parcelas, y Brugmansia sp. (trompeta de ángel), en la parcela 1 y 2, ambas consideradas las especies más destacadas. El índice de diversidad indicó una distribución homogénea en la composición florística, con pequeñas variaciones entre las parcelas.
Tabla 2
Riqueza, abundancia y diversidad
de plantas por parcela en Paraíso Quetzal Lodge (San Gerardo de Dota, Santa María, San José)
|
Especie de planta |
Parcela 1 |
Parcela 2 |
Parcela 3 |
Parcela 4 |
|
Digitalis purpurea (dedalera) |
0 |
1 |
1 |
0 |
|
Penstemon roseus (campanita rosa) |
2 |
0 |
0 |
0 |
|
Fuchsia spp. (fucsias) |
3 |
3 |
0 |
2 |
|
Brugmansia sp. (trompeta de ángel) |
2 |
2 |
0 |
0 |
|
Kniphofia sp. (lirio antorcha) |
0 |
0 |
0 |
2 |
|
Zantedeschia aethiopica (cala blanca) |
0 |
0 |
1 |
0 |
|
Myrcianthes storkii (guayabillo) |
0 |
0 |
1 |
1 |
|
Total de individuos |
7 |
6 |
3 |
5 |
|
Riqueza de especies |
3 |
3 |
3 |
3 |
|
Índice diversidad de Shannon |
1.079 |
1.011 |
1.099 |
1.055 |
Durante el muestreo se observó una variada composición florística destacándose especies de importancia ornamental y ecológica, pertenecientes a diferentes familias como Myrtaceae y Solanaceae, fueron las únicas familias presentes en ambos lugares.
Frecuencia de visitación de colibríes como indicador de interacción planta-polinizador en las parcelas
En la finca Doña Blanca, como se observa en la tabla 3, en la primera visita se registraron 89 interacciones correspondientes a cinco especies de colibríes; en la segunda visita, 74 interacciones correspondiente a cuatro especies; y en la tercera visita, 31 interacciones correspondientes también a cuatro especies. En total, durante las tres visitas de campo, se contabilizaron 194 individuos pertenecientes a seis especies de colibríes: Colibri cyanotus, Lampornis calolaemus, Lampornis castaneoventris, Panterpe insignis, Selasphorus flammula, y Selasphorus scintilla.
Cantidad de visitas por especies de colibríes en la finca Doña Blanca,
durante el mes de abril de 2024
|
|
Colibrí cyanotus |
Lampornis calolaemus |
Lampornis castaneoventris |
Panterpe insignis |
Selasphorus flammula |
Selasphorus scintilla |
Sin visita |
Total general |
|---|---|---|---|---|---|---|---|---|
|
Primera visita |
2 |
32 |
17 |
33 |
5 |
|
0 |
89 |
|
Agapanthus africanus (agapanto africano) |
|
|
|
|
|
|
0 |
0 |
|
Eriobotrya japónica (níspero) |
|
|
7 |
|
|
|
|
7 |
|
Psidium cattleianum (guayabita del Perú) |
|
|
|
|
|
|
0 |
0 |
|
Stachytarpheta mutabilis |
|
|
|
15 |
|
|
|
15 |
|
Strelitzia reginae (ave del paraíso) |
|
|
|
|
|
|
0 |
0 |
|
Streptosolen jamesonii (arbusto mermelada) |
2 |
32 |
10 |
16 |
5 |
|
|
65 |
|
Tibouchina urvilleana (arbusto de la gloria) |
|
|
|
2 |
|
|
|
2 |
|
Segunda visita |
|
19 |
8 |
39 |
8 |
|
0 |
74 |
|
Agapanthus africanus (agapanto africano) |
|
|
|
3 |
|
|
|
3 |
|
Eriobotrya japónica (níspero) |
|
|
2 |
1 |
|
|
|
3 |
|
Psidium cattleianum (guayabita del Perú) |
|
|
|
|
|
|
0 |
0 |
|
Stachytarpheta mutabilis (rabo de gato) |
|
|
|
11 |
|
|
|
11 |
|
Strelitzia reginae (ave del paraíso) |
|
|
|
1 |
|
|
|
1 |
|
Streptosolen jamesonii (arbusto mermelada) |
|
19 |
6 |
23 |
8 |
|
|
56 |
|
Tibouchina urvilleana (arbusto de la gloria) |
|
|
|
|
|
|
0 |
0 |
|
Tercera visita |
|
13 |
12 |
5 |
|
1 |
0 |
31 |
|
Agapanthus africanus (agapanto africano) |
|
|
|
|
|
|
0 |
0 |
|
Eriobotrya japónica |
|
2 |
|
|
|
|
|
2 |
|
Psidium cattleianum |
|
|
|
|
|
|
0 |
0 |
|
Stachytarpheta mutabilis |
|
|
|
|
|
|
0 |
0 |
|
Strelitzia reginae |
|
|
|
|
|
|
0 |
0 |
|
Streptosolen jamesonii |
|
10 |
12 |
5 |
|
1 |
|
28 |
|
Tibouchina urvilleana |
|
1 |
|
|
|
|
|
1 |
|
Total general |
2 |
64 |
37 |
77 |
13 |
1 |
0 |
194 |
Nota. Se muestra la cantidad de visitas registradas por cada especie de colibrí sobre cada especie de planta
durante las tres visitas de campo en la finca Doña Blanca, durante el mes de abril 2024.
En la finca Doña Blanca, la floración de Streptosolen jamesonii (arbusto mermelada) concentró la mayor parte de la actividad: de 149 visitas registradas sobre esta especie, Lampornis calolaemus aportó 40,9 %, Panterpe insignis aproximadamente un tercio (29,5 %) y Lampornis castaneoventris aportó 18,8 %. La participación de Selasphorus flammula fue más discreta (8,7 %), mientras que Colibri cyanotus (1,3 %) y Selasphorus scintilla (0,7 %) aparecieron de forma esporádica. A escala del sitio (n = 194), Stachytarpheta mutabilis (rabo de gato) concentró 13,4 % de las visitas, todas realizadas por P. insignis, lo que sugiere una asociación marcada con este recurso. En conjunto, S. jamesonii (76,8 % del total) y S. mutabilis se consolidan como las especies núcleo del jardín por su capacidad de atraer y sostener la interacción colibrí-planta.
Figura 3
Promedio de visitas de colibríes en parcelas para la finca Doña Blanca
en el mes de abril del 2024
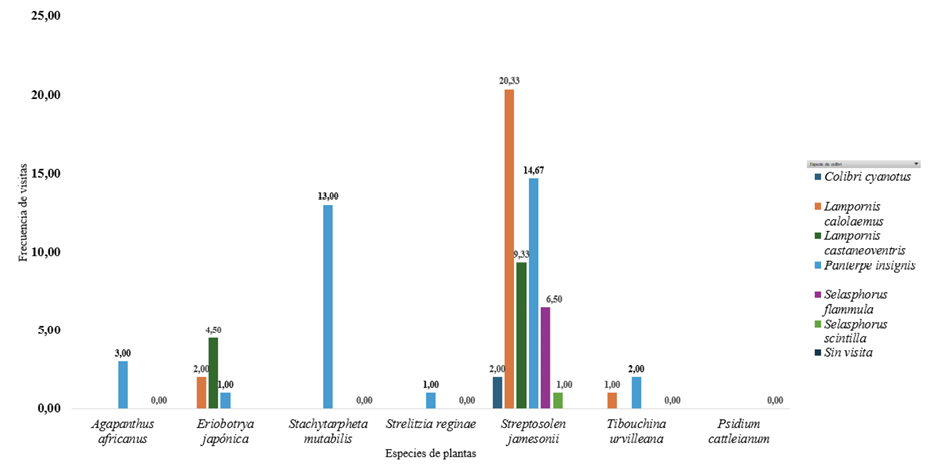
Nota. El gráfico muestra el promedio de interacciones entre colibrí y planta, según la frecuencia de observación
en las parcelas de la finca Doña Blanca.
De igual manera, se identificaron especies de colibríes con diferentes niveles de interacción: Selasphorus scintilla y Colibri cyanotus con la menor interacción, mientras Lampornis calolaemus y Panterpe insignis mostraron la mayor interacción en especies como Streptosolen jamesonii (arbusto mermelada). Por el contrario, Psidium cattleianum (guayabita del Perú) se clasificó como la especie de planta con nula interacción durante las visitas.
Se observaron diversos comportamientos de los colibríes: Panterpe insignis visitó con frecuencia el Streptosolen jamesonii (arbusto mermelada) para alimentarse. Por otro lado, Lampornis calolaemus y Lampornis castaneoventris utilizaron Eriobotrya japonica (níspero) únicamente como percha. En el caso de Agapanthus africanus (agapanto africano), las pocas interacciones registradas correspondieron también al uso del elemento vegetal como percha. Además, se observaron comportamientos territoriales, especialmente en Lampornis calolaemus.
En Paraíso Quetzal Lodge, la actividad se mantuvo estable entre las visitas, con 33, 34 y 27 interacciones registradas en la primera, segunda y tercera salida, respectivamente lo que representa 35,1 %, 36,2 % y 28,7 % del total, respectivamente, con una riqueza constante de cuatro especies de colibrí por visita (tabla 4). En el acumulado de la campaña (n = 94 interacciones), participaron seis especies, con un claro predominio de Panterpe insignis (45/94; 47,9 %), seguido por Eugenes spectabilis (31/94; 33,0 %); el 19,1 % restante se distribuyó entre las otras cuatro especies registradas. Este patrón sugiere una estructura de visitación centrada en dos taxones focales, consistente con un ensamblaje de alta montaña donde unas pocas especies concentran la mayor parte de la interacción.
Tabla 4
Cantidad de visitas por especies de colibríes en Paraíso Quetzal Lodge durante los meses
de mayo y junio de 2024
|
Colibrí cyanotus |
Eugenes spectabilis |
Lampornis castaneoventris |
Panterpe insignis |
Selasphorus flammula |
Selasphorus scintilla |
Sin visita |
Total general |
|||
|
Primera visita |
2 |
8 |
|
12 |
11 |
|
0 |
33 |
||
|
Brugmansia sp. (trompeta de ángel) |
2 |
3 |
|
|
|
|
|
5 |
||
|
Digitalis purpurea (dedalera) |
|
|
|
|
|
|
0 |
0 |
||
|
Fuchsia spp. (fucsias) |
|
4 |
|
4 |
9 |
|
|
17 |
||
|
Kniphofia sp. (lirio antorcha) |
|
1 |
|
|
2 |
|
|
3 |
||
|
Myrcianthes storkii (guayabillo) |
|
|
|
8 |
|
|
|
8 |
||
|
Penstemon roseus (campanita rosa) |
|
|
|
|
|
|
0 |
0 |
||
|
Zantedeschia aethiopica (cala blanca) |
|
|
|
|
|
|
0 |
0 |
||
|
Segunda visita |
1 |
17 |
2 |
14 |
|
|
0 |
34 |
||
|
Brugmansia sp. (trompeta de ángel) |
|
2 |
|
|
|
|
|
2 |
||
|
Digitalis purpurea (dedalera) |
|
|
|
|
|
|
0 |
0 |
||
|
Fuchsia spp. (fucsias) |
1 |
13 |
2 |
2 |
|
|
|
18 |
||
|
Kniphofia sp. (lirio antorcha) |
|
1 |
|
|
|
|
|
1 |
||
|
Myrcianthes storkii (guayabillo) |
|
1 |
|
12 |
|
|
|
13 |
||
|
Penstemon roseus (campanita rosa) |
|
|
|
|
|
|
0 |
0 |
||
|
Zantedeschia aethiopica (cala blanca) |
|
|
|
|
|
|
0 |
0 |
||
|
Tercera visita |
|
6 |
1 |
19 |
|
1 |
0 |
27 |
||
|
Brugmansia sp. (trompeta de ángel) |
|
|
|
3 |
|
|
|
3 |
||
|
Digitalis purpurea (dedalera) |
|
|
|
|
|
|
0 |
0 |
||
|
Fuchsia spp. (fucsias) |
|
3 |
1 |
1 |
|
1 |
|
6 |
||
|
Kniphofia sp. (lirio antorcha) |
|
3 |
|
|
|
|
|
3 |
||
|
Myrcianthes storkii (guayabillo) |
|
|
|
15 |
|
|
|
15 |
||
|
Penstemon roseus (campanita rosa) |
|
|
|
|
|
|
0 |
0 |
||
|
Zantedeschia aethiopica (cala blanca) |
|
|
|
|
|
|
0 |
0 |
||
|
Total general |
3 |
31 |
3 |
45 |
11 |
1 |
0 |
94 |
||
Nota. Esta tabla muestra la cantidad de visitas realizadas por especie de colibrí sobre cada especie de planta durante
las tres visitas de campo en Paraíso Quetzal Lodge.
En la figura 4, se resume el promedio de visitas por especie de colibrí y su distribución sobre la vegetación. Destaca Myrcianthes storkii (guayabillo) como el recurso más concurrido, impulsado por Panterpe insignis con un promedio de 11,67 visitas. A su vez, Fuchsia spp. (fucsias) se perfila como planta “nodo” del jardín, al recibir visitantes de las seis especies de colibrí registradas. Dentro de ese conjunto, resaltan los promedios de Selasphorus scintilla (9,00) y Eugenes spectabilis (6,67). En el extremo opuesto, Digitalis purpurea (dedalera), Zantedeschia aethiopica (cala blanca) y Penstemon roseus (campanita rosa) mostraron interacción nula durante el periodo de observación, lo que sugiere una relevancia limitada en el suministro de néctar en este contexto o una posible desincronización fenológica respecto a la actividad de los colibríes. En conjunto, el patrón confirma una estructura de uso del recurso con pocas plantas núcleo (Myrcianthes storkii y Fuchsia spp.) y varias especies con aportes marginales, aspecto clave para orientar el diseño y la priorización de especies en el jardín.
Figura 4
Promedio de visitas de colibríes en parcelas para Paraíso Quetzal Lodge durante los meses de mayo y junio del 2024
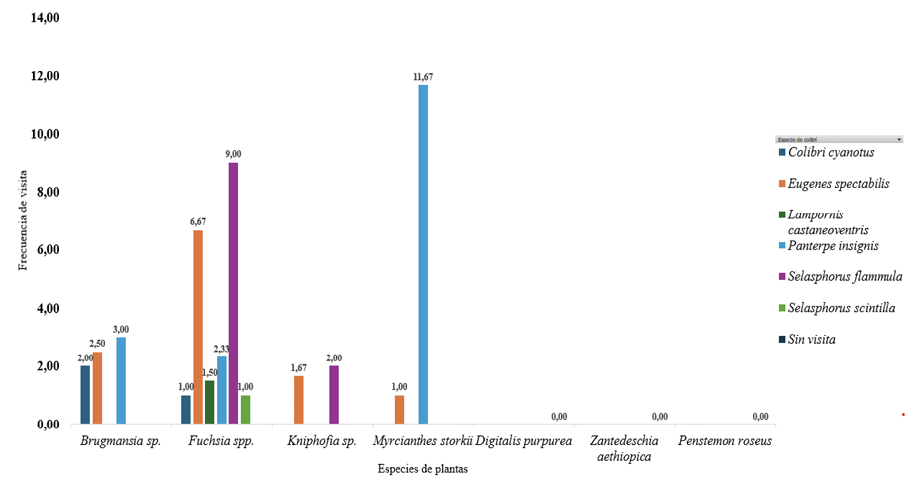
Nota. El gráfico muestra el promedio de interacciones colibrí- planta, según la frecuencia
de observación en las parcelas de Paraíso Quetzal Lodge.
Durante la segunda visita, se observó el acercamiento de Eugenes spectabilis en al menos cinco ocasiones cerca de una de las parcelas, alimentándose de residuos de papaya madura. Según lo indicado por el propietario, el comportamiento es inusual. Cabe resaltar que, aunque Selasphorus flammula y Selasphorus scintilla fueron observadas con mayor frecuencia en días fuera del periodo de muestreo, durante el presente estudio S. scintilla fue la especie con menor abundancia.
Plan de adecuación del jardín de observación de colibríes: diagnóstico de necesidades
La evidencia de campo indica que la visitación de colibríes se concentra en un conjunto reducido de especies con alta oferta de néctar y morfología compatible, mientras otras aportan poco o nada durante el periodo evaluado. En este contexto, Streptosolen jamesonii (arbusto mermelada) y Stachytarpheta mutabilis (rabo de gato) funcionan como plantas núcleo al sostener la mayor parte de las interacciones y estabilizar el flujo de visitas a lo largo del día. A su vez, Fuchsia spp. (fucsias) actúa como planta “nodo”, capaz de atraer simultáneamente a varias especies de colibríes, y Myrcianthes storkii (guayabillo) cumple un doble papel como recurso floral y, sobre todo, como percha o refugio que facilita el descanso, la defensa del parche y el retorno frecuente al néctar.
El desempeño del jardín mejora cuando esta composición se organiza en una arquitectura estratificada: herbáceas nectaríferas en el estrato bajo (p. ej., Streptosolen jamesonii –arbusto mermelada–, Stachytarpheta mutabilis –rabo de gato–); arbustos de soporte y floración intermedia (p. ej., Fuchsia spp. –fucsias–); y un componente arbóreo cercano (p. ej., Myrcianthes storkii –guayabillo–) que provee perchas sombreadas. Cuando las “islas de néctar” se ubican a corta distancia de las perchas, los colibríes reducen su costo de desplazamiento y prolongan su permanencia en el área de forrajeo, lo que se traduce en un mayor número de visitas observables y en una distribución más estable de la actividad.
En sentido contrario, varias especies ornamentales mostraron un desempeño marginal o nulo durante el muestreo (por ejemplo, Digitalis purpurea -dedalera-, Zantedeschia aethiopica -cala blanca-, Penstemon roseus -campanita rosa-, Agapanthus africanus -agapanto africano- o Strelitzia reginae -ave del paraíso-), ya sea por desacople fenológico con la ventana de observación o por incompatibilidad funcional con los visitantes registrados. En términos prácticos, conviene priorizar la instalación y el manejo de las especies que demostraron alta interacción (núcleo y nodo), agruparlas en bloques compactos e intercalar perchas inmediatas. Paralelamente, relevar o reubicar las especies de baja respuesta para optimizar el espacio, el mantenimiento y el rendimiento ecológico del jardín.
Discusión
Los patrones observados indican que la visitación de colibríes no se distribuye al azar, sino que se concentra en un subconjunto de especies con morfología floral compatible y oferta sostenida de néctar. Este resultado es coherente con la expectativa de compatibilidad pico–corola en sistemas colibrí–planta: las flores tubulares y de colores de alta detectabilidad (rojos/naranjas) tienden a recibir más visitas en ambientes montanos (Amaya et al., 2001; Fernández & Frías, 2021; Vargas et al., 2022). La mayor recurrencia sobre Streptosolen jamesonii (arbusto mermelada), Stachytarpheta mutabilis (rabo de gato), Fuchsia spp. (fucsias) y Myrcianthes storkii (guayabillo) se alinea con ese mecanismo, mientras que especies de corola abierta, como Psidium cattleianum (guayabita del Perú), mostraron baja o nula interacción, probablemente por favorecer a polinizadores generalistas (insectos) más que a colibríes (Amaya et al., 2001).
Las diferencias entre sitios sugieren el papel modulador de la arquitectura del jardín. En el espacio consolidado de Paraíso Quetzal, la coexistencia de estratos (herbáceas–arbustos–árboles) y la proximidad de perchas generaron escenarios de alimentación, defensa y retorno que sostienen interacciones repetidas. Por otra parte, en la finca Doña Blanca, con una estructura aún en desarrollo, la respuesta se concentró en unas pocas “plantas núcleo”. La disponibilidad inmediata de perchas es especialmente relevante porque reduce el costo energético del forrajeo y permite conductas de defensa del parche, lo que favorece una mayor permanencia en los puntos de recurso (Johnson et al., 2020; Bonifacio & Fredericksen, 2000) -figura 5-. La observación de forrajeo rutero (p. ej., Panterpe insignis) frente a forrajeo territorial (p. ej., Lampornis calolaemus) ayuda a explicar por qué ciertas plantas “nodo”, capaces de recibir varias especies, mantienen flujos de visita incluso bajo competencia interespecífica.
Figura 5
Especies de colibríes observados en el estudio durante los meses de abril, mayo
y junio del 2024

Nota. Especies de colibríes observadas en comportamiento de percha en Finca Doña Blanca y Paraíso Quetzal Lodge. (a) Selasphorus scintilla, (b) Panterpe insignis, (c) Colibri cyanotus y (d) Lampornis castaneoventris. Fotografías de Sergio Monge. Finca Doña Blanca, Jardín de Dota y Paraíso Quetzal Lodge, San Gerardo de Dota, 2024.
Un elemento adicional es la fenología, entendida como la relación entre los fenómenos biológicos y los cambios estacionales. La actividad registrada entre abril y junio sugiere que la continuidad de floración de las especies clave favoreció la estabilidad del recurso durante el periodo de muestreo, mientras que otras plantas ornamentales permanecieron desacopladas, ya sea por asincronía fenológica o por limitaciones morfológicas. Este hallazgo coincide con la idea de que los colibríes ajustan su dieta según la disponibilidad de recursos, manteniendo el equilibrio del sistema sin requerir una especialización estricta (Cardozo et al., 2018). La conducta oportunista de Eugenes spectabilis, alimentándose de papaya fuera de las parcelas, refuerza esta flexibilidad nutricional, sin implicar necesariamente escasez de néctar, pero sí la capacidad de incorporar recursos no florales cuando están disponibles.
Estos resultados respaldan, en conjunto, las siguientes hipótesis: (i) la compatibilidad morfológica entre flores y colibríes (Amaya et al., 2001; Fernández & Frías, 2021; Vargas et al., 2022); (ii) el efecto de perchas y microhábitats de descanso sobre la permanencia y la intensidad de uso de los recursos (Johnson et al., 2020; Bonifacio & Fredericksen, 2000); y (iii) el beneficio de la estratificación estructural del jardín para sostener interacciones estables. Desde una perspectiva territorial, la posibilidad de diseñar arreglos que hagan visibles estas dinámicas y de incorporarlas en programas de educación ambiental, turismo de observación y apropiación del paisaje apunta a generar sinergias entre conservación y desarrollo local en Los Santos (MINAE, 2021; González, 2018).
En cuanto a las limitaciones y proyecciones, la inferencia del estudio se circunscribe a una ventana temporal específica (abril–junio) y a un esfuerzo basado en conteos directos. En investigaciones futuras, convendría convertir los registros a tasas de visita (visitas/10 min) e incorporar una covariable de disponibilidad floral (número de flores abiertas o inflorescencias por unidad de área) para diferenciar la abundancia de la preferencia de los colibríes. La aplicación de modelos de conteo (GLMM Poisson o NegBin con offset por esfuerzo) y de métricas de redes ecológicas (por ejemplo, H2´, NODF) permitiría cuantificar la especialización, el anidamiento y los efectos de sitio o especie de planta sobre las interacciones. Asimismo, un monitoreo estacional que contemple periodos lluviosos y secos permitiría evaluar la estabilidad fenológica y la plasticidad trófica de los colibríes a lo largo del año.
La evidencia obtenida sugiere que, en el diseño y manejo de jardines polinizadores se debe priorizar la conformación de un núcleo nectarífero compuesto por herbáceas y arbustos de corola tubular, acompañado de perchas próximas (arbustos/árboles). Además, una adecuada estratificación vegetal puede reducir las distancias de vuelo. Esta estructura maximiza la atracción y retención de las aves, y ofrece un modelo replicable para jardines de alta montaña, con potencial para integrarse a rutas de observación y actividades de educación ambiental en la región (MINAE, 2021; González, 2018).
Conclusiones y recomendaciones
Las observaciones realizadas en la finca Doña Blanca y Paraíso Quetzal confirman que la interacción colibrí–planta no es homogénea, sino que se concentra en un conjunto reducido de especies con morfología compatible y oferta sostenida de néctar (véanse figuras 3 y 4 y tabla 4). En Doña Blanca, el flujo de visitas se articuló alrededor de plantas núcleo como Streptosolen jamesonii (arbusto mermelada) y Stachytarpheta mutabilis (rabo de gato), mientras que en Paraíso Quetzal el uso recurrente de Myrcianthes storkii (guayabillo) y el papel “nodo” de Fuchsia spp. (fucsias) sostuvieron la actividad de múltiples especies de colibrí. En contraste, taxones como Psidium cattleianum y Penstemon roseus aportaron poco o nada durante el periodo evaluado, evidenciando que no toda especie ornamental es funcional para la interacción.
Más allá de la composición, la estructura del jardín resultó determinante. La proximidad entre “islas de néctar” y perchas (arbustos/árboles) favoreció conductas de descanso, defensa y retorno al recurso, incrementando la permanencia de las aves en los parches más productivos. En conjunto, los patrones observados respaldan las hipótesis de la compatibilidad morfológica, el efecto de perchas y la estratificación vertical como motores de la visitación, y explican la diferencia entre un sitio consolidado (Paraíso Quetzal) y otro en consolidación (Doña Blanca) en términos de diseño y desempeño.
En síntesis, el estudio demuestra que pocas especies altamente efectivas, integradas en una arquitectura vegetal estratificada con perchas próximas, maximizan la atracción y la retención de colibríes. Este resultado ofrece una base empírica directa para orientar la composición y el arreglo de jardines de observación en ambientes de alta montaña.
Se recomienda centrar el diseño en dos o tres especies núcleo de floración sostenida (p. ej., Streptosolen jamesonii -arbusto mermelada-, Stachytarpheta mutabilis -rabo de gato- y Fuchsia spp. -Fucsias-) complementadas con un elemento de percha inmediato (p. ej., Myrcianthes storkii -guayabillo-). Estas especies deben agruparse en bloques compactos e intercalarse con perchas a distancias cortas para reducir el costo de desplazamiento y limitar la exclusión por territorialidad. En los casos donde se detecten especies con desempeño marginal o nulo, se recomienda relevarlas o reubicarlas para optimizar espacio y mantenimiento, priorizando la continuidad de floración mediante poda y siembras escalonadas.
Se sugiere establecer un monitoreo estacional (al menos durante las estaciones lluviosa y seca) mediante métricas comparables —por ejemplo, visitas por ١٠ minutos por especie y parcela, flores abiertas/m² y disponibilidad de perchas—, de modo que el jardín pueda ajustarse adaptativamente según resultados. En Doña Blanca, esto implica reforzar el núcleo nectarífero y aumentar puntos de percha próximos; en Paraíso Quetzal, mantener el mosaico que distribuye la presión de forrajeo y gestionar los cuellos de botella en senderos para reducir la dominancia de una o dos especies de colibrí. Con este esquema, el jardín no solo mejorará su valor ecológico, sino también su potencial de educación ambiental y aviturismo a escala local.
Agradecimientos
Se agradece al Laboratorio de Investigación de Agua y Suelos de San Marcos por el acceso a sus instalaciones y el apoyo en la recolección de datos. Se expresa gratitud a los propietarios de Paraíso Quetzal Lodge y de la finca Doña Blanca por permitir la realización del estudio en sus áreas. Asimismo, se agradece a los profesionales Paola Brenes Rojas y Sergio Monge por su valioso acompañamiento y aportes durante las etapas de observación y desarrollo del estudio.
Referencias
Amaya, M., Stiles Hurd, F. G., y Rangel Churio, J. O. (2001). Interacción planta-colibrí en Amacayacu (Amazonas, Colombia): una perspectiva palinológica. Caldasia, 23(1), 301–322.
https://revistas.unal.edu.co/index.php/cal/article/view/17680
Audesirk, T., Audesirk, G., y Byers, B. E. (2014). Biología general. Pearson Educación.
Brenes Rojas, P., Vargas Meneses, C., & Muñoz Picado, A. (2024). Guía metodológica para el desarrollo de jardines de observación de colibríes para la zona de Los Santos, San José, Costa Rica, 2024. Revista Biocenosis, 35(2), 17–33.
https://doi.org/10.22458/rb.v35i2.5601
Burbano-Álvarez, W. (2013). Morphological co-evolution between hummingbirds´ peaks and Plants´ Corollas? Biocyt, 6(20), 398-409. https://biblat.unam.mx/hevila/Biocytbiologiacienciaytecnologia/2013/vol6/3.pdf
Bonifacio, M. y Fredericksen, T. (2000). Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal. Proyecto de Manejo Forestal BOLFOR.
http://www.bio-nica.info/biblioteca/mostacedo2000ecologiavegetal.pdf
Cardozo, J. S.P., Fajardo, D.M., Torres, S. J.V. y Sepúlveda, M. D. P. N. (2018). Análisis de la Interacción planta-colibrí en la reserva Mirador Recreo: una perspectiva coevolutiva. Revista de la Asociación Colombiana de Ciencias Biológicas, 1(30), 114-120.
https://revistaaccb.org/r/index.php/accb/article/view/158
Faigón, M. (2021). Causas y consecuencias de la disminución de polinizadores silvestres. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Fernández, N., Frías, A. (2021). Caracterización de recursos florales aprovechados por colibríes en una gradiente de disturbio en bosques montanos del cantón de Cuenca. [Trabajo Final de Graduación, Universidad del Azuay]. https://dspace.uazuay.edu.ec/bitstream/datos/11226/1/16763.pdf
González M, A. (2018). Distribución y riqueza espacial de colibríes con relación a coberturas y uso del suelo en la Región Andina. [Trabajo de grado, Universidad De La Salle]. https://ciencia.lasalle.edu.co/server/api/core/bitstreams/e314732f-a8ff-44dd-838e-26f703410050/content
Johnson, D.S., Kiepiel, I., & Robertson, W.A. (2020). Functional consequences of flower curvature, orientation and perch position for nectar feeding by sunbirds. Biological Journal of the Linnean Society, 131(4), 822–834.
https://doi.org/10.1093/biolinnean/blaa154
Ministerio de Ambiente y Energía, Sistema Nacional de Áreas de Conservación., y Agencia de Cooperación del Gobierno de Alemania. (2021). Guía y catálogo de Soluciones basadas en la naturaleza. https://biocorredores.org/biodiver-city-sanjose/sites/default/files/2021-10/05-jard%C3%ADn%20de%20polinizaci%C3%B3n%20DIGITAL.PDF
Plataforma Intergubernamental Científico Normativa sobre Diversidad Biológica y Servicios de los Ecosistemas. (2016). Resumen para los responsables de formular políticas del informe de evaluación de la Plataforma Intergubernamental Científico-normativa sobre Diversidad Biológica y Servicios de los Ecosistemas sobre polinizadores, polinización y producción de alimentos. https://files.ipbes.net/ipbes-web-prod-public-files/2020-02/ipbes_global_assessment_report_summary_for_policymakers_es.pdf
Rodríguez Fernández, C. I., Stiles, F. G. (2005). Análisis ecomorfológico de una comunidad de colibríes ermitaños (Trochilidae, Phaethorninae) y sus flores en la Amazonia colombiana. Ornitología Colombiana, (3), 7-27. https://pdfs.semanticscholar.org/3f9e/fcf23f158583d9fc51b314b9b86c71576e77.pdf
Vargas Valverde, I., Campos Alvarado, A., Niño Rodríguez, N., Randy, S., Piñanez Espejo, Y., Hernández Rivera, A. y Avalos, G. (2022). Redes de interacción colibrí-planta en un área abierta con robledales en el Cerro de La Muerte, Costa Rica. Zeledonia 26 (1), 26-38.
https://www.zeledonia.com/uploads/7/0/1/0/70104897/zeledonia_26.1_2022.pdf
1 Estudiante Manejo de Recursos Naturales. Universidad Estatal a Distancia (UNED), Laboratorio de Investigación
en Aguas y Suelos, LIAS – San Marcos, Costa Rica. rebecanavarrof29@gmail.com2 Ingeniera Agrónoma. Universidad Estatal a Distancia (UNED), Laboratorio de Investigación en Aguas y Suelos, LIAS - San Marcos, Costa Rica. Investigadora. pbrenes@uned.ac.cr
3 Ingeniera civil. Universidad Estatal a Distancia (UNED), Laboratorio de Investigación en Aguas y Suelos, LIAS - San Marcos, Costa Rica. Coordinadora. cvargasm@uned.ac.cr
